Supplement
to a paper published
in Castanea,
Vol. 66(1-2):154–205, March/June 2001
Musical selection: Les
Barricades
Mysterieuses by François
Couperin (1668-1733)
All photography (except as noted) by James R. Allison and Copyright 2010. All
rights reserved.

NOTE: AT THE BOTTOM OF THIS
PAGE IS A
LINK TO A SPECIAL WEB VERSION OF THE ORIGINAL PAPER!
Vascular Flora
of Ketona Dolomite Outcrops in Bibb County, Alabama
by
JAMES
R. ALLISON*
and
TIMOTHY
E. STEVENS**
*P.O. Box 511
Rutledge,
Georgia 30663
**Alabama
Department of Public Health Laboratories
8140 AUM Dr.,
Montgomery,
Alabama 36124
ABSTRACT
Explorations
since
1992 in Bibb County, Alabama, have revealed an extraordinary,
undescribed
glade community
developed over the Ketona Formation, an
unusually pure
dolomite. Eight new endemic taxa were found: Castilleja
kraliana,
Coreopsis grandiflora var. inclinata,
Dalea
cahaba,
Erigeron strigosus var. dolomiticola,
Liatris
oligocephala,
Onosmodium
decipiens,
Silphium
glutinosum,
and Spigelia gentianoides var. alabamensis.
In
assessing systematic relationships of the Erigeron
and
Silphium,
two additional undescribed taxa, not of Bibb County,
were discerned, E. strigosus var. calcicola
and S. perplexum.
Seven state records were discovered: Solanum pumilum,
last collected
in 1837 and presumed extinct; Astrolepis
integerrima,
disjunct from
Texas; Paronychia
virginica,
bridging a gap between Arkansas and
Virginia; Baptisia australis var. australis,
Rhynchospora
capillacea,
R.
thornei
and Spiranthes
lucida.
More than 60 plant taxa
of conservation concern occur on or near these glades, marking them as
one of the most significant reservoirs of botanical diversity in the
eastern
United States.
INTRODUCTION
Kenneth J. Wurdack reports
about an historic visit to
Browne's
Dam, site of
major Ketona dolomite glades on both sides of the Little Cahaba River,
in "The Lectotypification and 19th Century
History of Croton alabamensis (Euphorbiaceae
s.s.)" (Sida 22: 469 – 483. 2006) [available at http://www.andestoamazon.com/Sida/PDF/PDF22(1)/24_Wurdack_Croton_469-483.pdf].
Century
History of Croton alabamensis (Euphorbiaceae
s.s.)" (Sida 22: 469 – 483. 2006) [available at http://www.andestoamazon.com/Sida/PDF/PDF22(1)/24_Wurdack_Croton_469-483.pdf].
Referring to Eugene Allen Smith
(1841–1927), appointed Alabama's State Geologist in 1873, Wurdack states (p. 472):
Smith
first explored Bibb County in 1873 and in more detail in 1875. On August 17,
1875 he collected ore samples and explored associated Brighthope ironworks or
“bloomery” (originally called Little Cahaba Furnace and in operation by
1850; Ellison 1984*), the first blast furnace in the county and about 140 m
above a wooden dam (Browne’s Dam) on the Little Cahaba River that provided its
power. The seats for the dam timbers appear today as a series of steps cut into
a riverbank rock outcrop at the Bibb County Glades Preserve near Bulldog Bend
(pers. observ; Ellison 1993:51**). The apparent present day natural state of
this area, which contains some of the finest Ketona Glades, is remarkable
considering the past destructive activities engendered by the adjacent
ironworks, although it is also possible that disturbance supported or enlarged
the glade community.
*
ELLISON, R.C. 1984. Bibb County
Alabama. The first hundred years, 1818-1918. The University of Alabama Press.
**
ELLISON, R.C. 1993. Place names of
Bibb County: Abercrombie to Zuzu. Cahaba Trace Commission, Brierfield, Alabama.
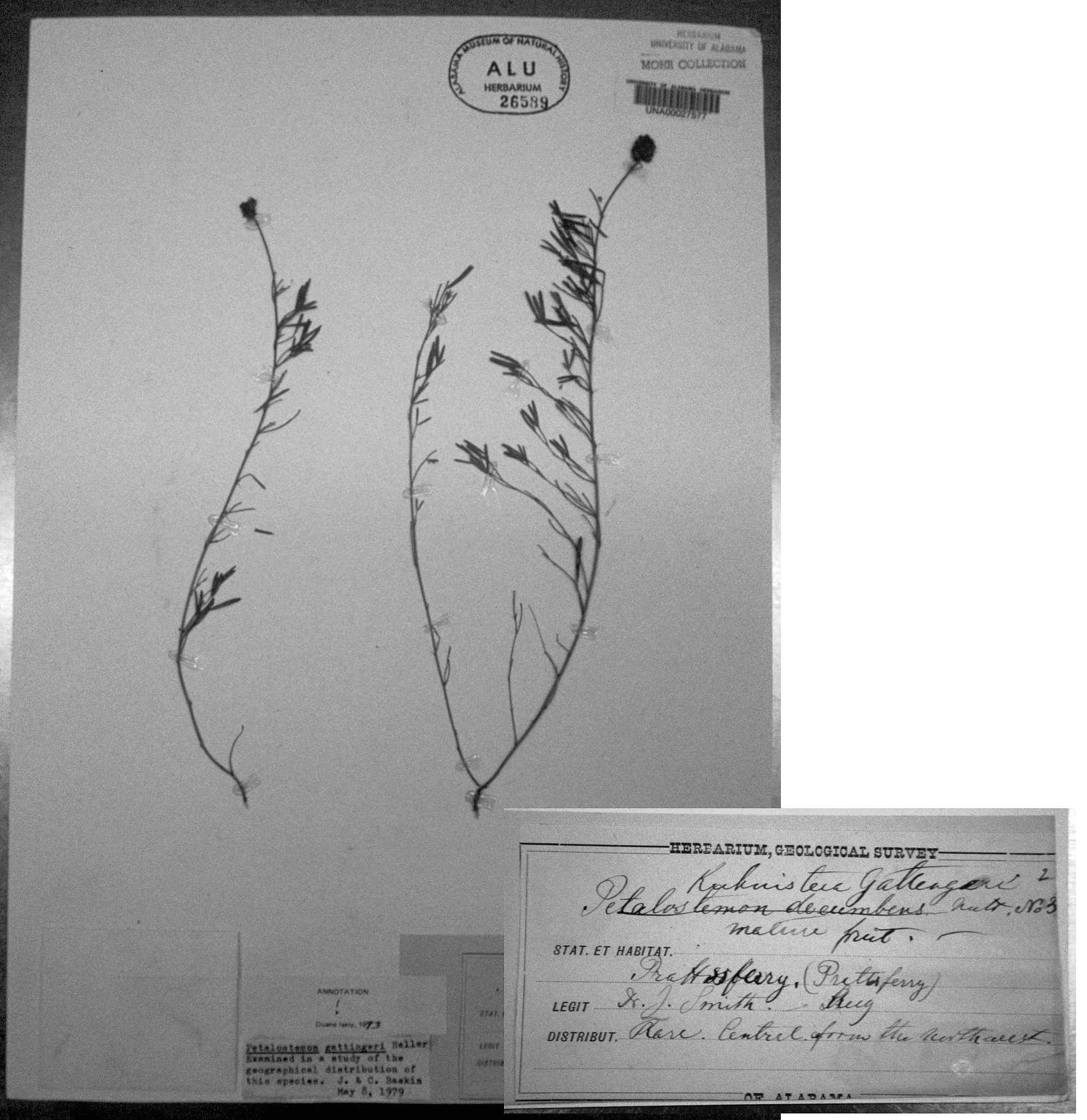
This would seem to fix the date of Smith's collection of Dalea cahaba
(at right; referenced in the Castanea paper on p. 168) as August 17,
1875; the label merely states "August." The locality on Smith's
label, "Pratts Ferry" (known to current Bibb County residents as River
Bend) must be interpreted as a very general locality. The collection undoubtedly
came from Browne's Dam, based on Wurdack 2006.
Although according to Wurdack, Charles Mohr (1824-1901)
visited Bibb County on two occasions, no collections by Mohr are known of any of
the Ketona glade endemics. Mohr's 1882 trip was focused on getting material of
the yet-to-be-described Croton alabamensis E. A. Sm. ex Chapm., when he reportedly explored
an area of lower Six Mile Creek with success. Today there are a few small glades
in this vicinity that support the endemic Erigeron, Onosmodium,
and Silphium. Mohr probably saw one or more of the small glades at and
near the mouth of Six Mile Creek, after following it down to the Little Cahaba
River. Mohr must have reached the river because in his Plant Life of Alabama
(p. 93) he reports finding Quercus breviloba (Torr.) Sarg. [actually the
not-yet-described Q. austrina Small] "on the limestone
hills lining the Little Cahaba River in 1882." However, as Mohr's 1882 trip
was in mid-November, any of the endemics would likely have gone unnoticed.
Wurdack reports that Mohr returned to Bibb County the following June, again in
search of the Croton, but only mentions Pratt's Ferry as a destination.
This would have been a prime time to collect some Ketona glade plants, but the
nearest known glade to that location is 11 km away. The one endemic that strays
a bit from the glades, Silphium glutinosum, is found at the base of a
limestone bluff not far upstream from Pratt's Ferry. If Mohr saw the Silphium
in June (1883) it would likely not yet have been in flower.
BIOLOGICAL COMMUNITIES
Among the species listed in the Castanea paper as characteristic of partly
shaded glade-forest ecotones or islands of woody vegetation on the open glades
(also mentioned later among the non-endemic rarities) was Carex eburnea Boott.
Study of material Emily L. Gillespie collected from the Bibb County
Glades was helpful to her in molecular and quantitative morphological analyses
she conducted that indicated that the closely related C. mckittrickensis
Ball should be reduced to synonymy under C. eburnea [Phylogeography of Carex
eburnea (Cyperaceae) and the Systematics of the Carex eburnea
Complex. 2005. MS thesis, Appalachian State Univ., Boone, NC (available at http://www.biology.appstate.edu/herbarium/Research/GILLESPIE%20thesis.pdf)]. Her overall data
set supported hypotheses that "the northwestern-most populations of C.
eburnea are basal, and that populations in the south and east are
derived," and that "the ancestor to the C. eburnea [complex]
migrated from Asia into North America via the Bering Land Bridge."
I confess to feeling "ambushed" at one point when I read
"Xeric Limestone Prairies of Eastern United States: Review and Synthesis
(Botanical Review 72: 235-272. 2006), by Patrick J. Lawless, Jerry M. Baskin,
and Carol C. Baskin.
In the Castanea paper about the Bibb County Glades, I wrote in the last paragraph under
Biological Communities that
Baskin et al. (1994*)
attempted to resolve inconsistencies in the use of such terms as
"glades," "barrens," and "limestone prairies" that
have been used in discussing openings, dominated by grasses and forbs, that are
developed over calcareous bedrock. They devised over a dozen criteria useful for
assigning such places to one of three general categories: limestone glade, xeric
limestone prairie, or barrens. The Ketona Glades fail several to many criteria
for each of their three categories, but come closest to the "xeric
limestone prairie" class. Since they differ from limestone prairies by
developing over dolomite rather than limestone, by containing multiple endemics,
and by supporting two species of Leavenworthia, the simplest course
would be to establish a fourth category to accommodate the Ketona Glades.
*
Baskin, J.M., C.C. Baskin, and E.W. Chester. 1994. The Big Barrens Region of
Kentucky and Tennessee: further observations and considerations. Castanea
59:226-254.
Lawless et al. (2006) was derived from Lawless' dissertation (available at http://archive.uky.edu/bitstream/10225/141/Lawless.pdf).
After quoting the paragraph given above from the Ketona Glades paper almost
verbatim, the following "criticisms" occur in both texts with only
trivial differences (quoted here from Lawless et al 2006):
However, as
discussed above, XLPs [xeric limestone prairies] occur on dolomite in various regions throughout the
geographic range of this community type in the eastern United States.
Furthermore, XLP endemics (Table IV) occur in regions other than the Cahaba
River valley in Alabama, particularly in the Ozark Plateaus in Missouri and
Arkansas. In addition, Leavenworthia spp., some of which primarily are limestone
cedar glade endemics (e.g., Leavenworthia alabamica, Leavenworthia
exigua var. exigua, and Leavenworthia exigua var. laciniata),
occur in other XLPs in the eastern United States (Baskin & Baskin, 1977;
Maxwell, 1987; DeSelm, 1988, 1991, 1993; DeSelm & Chester, 1993; Ver Hoef et
al., 1993; Webb et al., 1997; Gardner & Minnie, 2004; Lawless et al., 2004).
One unacquainted with
the literature would likely conclude from the preceding language that some
oversights had been committed in regard to this matter in the Castanea paper.
However the reality is that, in his 2005 dissertation (several years after the
paper on the Ketona Glades) Lawless had significantly redefined the meaning of
the term "xeric limestone prairie" from that employed in Baskin et al.
(1994). His redefinitions included the unfortunate extension of the
community-type to include expressions developed over dolomite without
changing the name to xeric calcareous prairie, as logic and
scientific clarity would seem to dictate. This resulted in his classifying all
the Bibb County endemics and other species found in his redefined community-type
as xeric limestone prairie endemics and the Leavenworthia species
as cedar glade endemics co-occurring on XLPs. However, according to Baskin et
al. 1994 (p. 240), limestone cedar glades contain endemic taxa while xeric limestone
prairies do not. They also characterized Leavenworthia (p. 248) as "a genus
endemic to cedar glades."
Finally, do the
Ketona Dolomite Glades truly fit this community-type, as redefined by Lawless,
Baskin, and Baskin, even if the name is corrected to xeric calcareous
prairie? Or, as suggested in our 2001 paper, with their rich suite of
endemics are the Ketona dolomite glades of Bibb County, Alabama different enough to be made a category of their own?
Interestingly, in Baskin et al. (1994) limestone cedar glades were characterized
as being a natural, edaphic climax, while xeric limestone prairies were said to
be an anthropogenic disclimax, which would certainly explain their
lack of endemics. They did concede that a few XLPs may be natural. Given this,
it seems telling to me that Lawless (2005) stated (p. 178) that "In
summary, XLPs of eastern United States are either edaphic climaxes (Ketona
Dolomite sites) or subclimaxes (sensu Weaver and Clements, 1938) that originated
from and are perpetuated by anthropogenic disturbance (burning, cutting, and/or
agricultural practices)." The answer to my question that began this
paragraph seems an obvious "No!"
NOTEWORTHY
ELEMENTS OF THE FLORA:
ENDEMIC
TAXA
Castilleja kraliana J. Allison,
named for Robert Kral, Professor Emeritus
of Botany, Vanderbilt University. Click on the scientific name to see
an image
of a flowering individual (with blue flowers of Amsonia
ciliata Walt.
var. tenuifolia
(Raf.) Woods. in the background).
 Castilleja
coccinea (L.) Spreng., Indian paintbrush (left), is the best known—almost
the
only— representative in
the
states east of the Mississippi Rive
Castilleja
coccinea (L.) Spreng., Indian paintbrush (left), is the best known—almost
the
only— representative in
the
states east of the Mississippi Rive r of a genus that is
quite diverse
in the western states, Mexico and South
America. It is
a fairly typical Castilleja
in that most of the coloration
of the flower comes not from the corolla but from a deeply lobed,
subtending
bract and from the apical, lobed portion of the calyx. It is less
typical
in that it is an annual (or sometimes biennial), while the majority of
species are perennial, some of them even shrubs. Even more unusual is
the
degree of fusion of the calyx lobes. In the great majority of Castilleja
species there are four
well developed calyx lobes, but in
C. coccinea
fusion of the lobes
along the lateral clefts has resulted in a two-lobed calyx, (the only
vestige
of the lateral clefts of its ancestral form is that the two remaining
lobes may
each have a
very shallow, apical indentation). In Alabama, at least, C.
coccinea
has
been reported only where the soil reaction is acidic, growing over
sandstone. The
Castilleja
of the alkaline soils of the Ketona Glades (image at right, not to same
scale as preceding image) is clearly closely related to C.
coccinea,
by virtue of its annual (or biennial) duration and, more importantly,
its similar calyx morphology (image shows a flower with bract
r of a genus that is
quite diverse
in the western states, Mexico and South
America. It is
a fairly typical Castilleja
in that most of the coloration
of the flower comes not from the corolla but from a deeply lobed,
subtending
bract and from the apical, lobed portion of the calyx. It is less
typical
in that it is an annual (or sometimes biennial), while the majority of
species are perennial, some of them even shrubs. Even more unusual is
the
degree of fusion of the calyx lobes. In the great majority of Castilleja
species there are four
well developed calyx lobes, but in
C. coccinea
fusion of the lobes
along the lateral clefts has resulted in a two-lobed calyx, (the only
vestige
of the lateral clefts of its ancestral form is that the two remaining
lobes may
each have a
very shallow, apical indentation). In Alabama, at least, C.
coccinea
has
been reported only where the soil reaction is acidic, growing over
sandstone. The
Castilleja
of the alkaline soils of the Ketona Glades (image at right, not to same
scale as preceding image) is clearly closely related to C.
coccinea,
by virtue of its annual (or biennial) duration and, more importantly,
its similar calyx morphology (image shows a flower with bract  manually
deflexed the better to show the calyx, which I also spread in order to
show more of the corolla). Castilleja
kraliana's bracts and
calyx lobes, however, are normally
bright yellow, while those of C.
coccinea are usually red (coccinea
is Latin for "scarlet"). Rare yellow forms of the latter, however, have
long
been known to occur
(image at left is a view from above), so the yellow coloration of the
flowers of C.
kraliana is not
manually
deflexed the better to show the calyx, which I also spread in order to
show more of the corolla). Castilleja
kraliana's bracts and
calyx lobes, however, are normally
bright yellow, while those of C.
coccinea are usually red (coccinea
is Latin for "scarlet"). Rare yellow forms of the latter, however, have
long
been known to occur
(image at left is a view from above), so the yellow coloration of the
flowers of C.
kraliana is not definitive in
and of itself. However, the bracts of
C. kraliana
are also usually entire (as
is apparent in the image above, right), while those of C.
coccinea,
whatever their color, are always deeply cleft and in vigorous plants,
the main
lobes are often themselves lobed. Occasionally the bracts
of C. kraliana
are shallowly lobed at
anthesis; after anthesis the bracts undergo a transformation, becoming
larger and more leaflike (greening, and any lobing becoming much more
prominent).
Were it not for the consistently smaller flowers of C.
kraliana,
I might have chosen to make it a subspecies of C.
coccinea. The
yellow coloration and smaller flower size of C.
kraliana are
apparently
adaptations promoting pollination by bumblebees (image at right),
rather
than hummingbirds, the chief pollinators of C.
coccinea (of the usual,
red forms, anyway) .
definitive in
and of itself. However, the bracts of
C. kraliana
are also usually entire (as
is apparent in the image above, right), while those of C.
coccinea,
whatever their color, are always deeply cleft and in vigorous plants,
the main
lobes are often themselves lobed. Occasionally the bracts
of C. kraliana
are shallowly lobed at
anthesis; after anthesis the bracts undergo a transformation, becoming
larger and more leaflike (greening, and any lobing becoming much more
prominent).
Were it not for the consistently smaller flowers of C.
kraliana,
I might have chosen to make it a subspecies of C.
coccinea. The
yellow coloration and smaller flower size of C.
kraliana are
apparently
adaptations promoting pollination by bumblebees (image at right),
rather
than hummingbirds, the chief pollinators of C.
coccinea (of the usual,
red forms, anyway) .
 At
left is an image that indicates the relative flower sizes of the two
species;
it was created by placing pressed Castilleja
specimens on a
flat
bed scanner. In it, four inflorescences of the smaller-flowered C.
kraliana
are flanked by one inflorescence each of yellow- and red-flowered C.
coccinea. Left to right [all
US]: Allison 10478,
Izard
County,
Arkansas, 19 April 1999; Allison
10466
[an isotype], Bibb County, Alabama, 15
April
At
left is an image that indicates the relative flower sizes of the two
species;
it was created by placing pressed Castilleja
specimens on a
flat
bed scanner. In it, four inflorescences of the smaller-flowered C.
kraliana
are flanked by one inflorescence each of yellow- and red-flowered C.
coccinea. Left to right [all
US]: Allison 10478,
Izard
County,
Arkansas, 19 April 1999; Allison
10466
[an isotype], Bibb County, Alabama, 15
April 1999; Allison 8270,
Lumpkin County, Georgia, 8 May 1994.
The
difference in flower size would be even more apparent in fresh material.
1999; Allison 8270,
Lumpkin County, Georgia, 8 May 1994.
The
difference in flower size would be even more apparent in fresh material.
As discussed in
the Castanea
paper, at the westernmost glade where Castilleja
kraliana occurs, "Eastside
Glade" (about 0.3 km east of the Cahaba
River), Tim Stevens and I found that some of the plants had bracts and
calyces orangish-tinged (image at right), and that some had slightly
more
deeply lobed bracts than is usual for the species (but note that the
plant
shown has the entire bracts usual in the species, as the anomalous
character-states
were not correlated). Perhaps one or more
undetected populations of the widespread C.
coccinea occur or
occurred
within the valley of the Cahaba River, and a past hybridization event
resulted
in the infusion of some C.
coccinea alleles into the
comparatively
isolated population of C.
kraliana closest to the river.
The
difference
in pollinators, however would seem to make this unlikely. Another
hypothesis
is that C. coccinea
was once an element of the flora of the
Ketona
Glades, and the process of its replacement by C.
kraliana is
virtually,
but not absolutely, complete.
Coreopsis grandiflora
Hogg ex Sweet var. inclinata
J. Allison. Inclinata,
the Latin word for "leaning,"
refers to the unusual, leaning
habit
of this variety. Click on the scientific name to see an image of a
flowering
stem, with the Little Cahaba River in the background.
 The
photo at left shows the
typical
habitat of Coreopsis
grandiflora var. inclinata.
It
grows in shallow soil over Ketona Dolomite, where it is
The
photo at left shows the
typical
habitat of Coreopsis
grandiflora var. inclinata.
It
grows in shallow soil over Ketona Dolomite, where it is subjected to
little
competition, either with other species or with other individuals of its
own kind (note the scattered occurrence of the yellow flowers). Its low
stature, due to the leaning habit, is also readily apparent in this
glade
scene. The other varieties are, by contrast, erect plants, and more
gregarious
as well (as shown in the picture of var. grandiflora
at right,
taken
on a Bibb County roadside). There the erect form has adaptive value in
competing
for light, both for photosynthesis and for enhanced visibility of
flowers to
pollinators. Even though the heads of var. inclinata
are
usually borne
within 2 dm of the substrate, the sparseness of taller vegetation
leaves the
flowers well exposed and easily perceived by the visitor, whether human
or
insect. Given the severely drought-prone habitat where it is found, the
adaptive
value of var. inclinata’s
peculiar habit, at least to an
insect-pollinated
species, is clear: a plant that is able to grow low to the ground and
yet still
attract pollinators can subsist with less moisture than an erect form
more
exposed to the drying effect of winds.
subjected to
little
competition, either with other species or with other individuals of its
own kind (note the scattered occurrence of the yellow flowers). Its low
stature, due to the leaning habit, is also readily apparent in this
glade
scene. The other varieties are, by contrast, erect plants, and more
gregarious
as well (as shown in the picture of var. grandiflora
at right,
taken
on a Bibb County roadside). There the erect form has adaptive value in
competing
for light, both for photosynthesis and for enhanced visibility of
flowers to
pollinators. Even though the heads of var. inclinata
are
usually borne
within 2 dm of the substrate, the sparseness of taller vegetation
leaves the
flowers well exposed and easily perceived by the visitor, whether human
or
insect. Given the severely drought-prone habitat where it is found, the
adaptive
value of var. inclinata’s
peculiar habit, at least to an
insect-pollinated
species, is clear: a plant that is able to grow low to the ground and
yet still
attract pollinators can subsist with less moisture than an erect form
more
exposed to the drying effect of winds.
The
black and white image at lower right is of representative
pressed specimens [UNA] of Coreopsis
grandiflora, all collected
24–26 April
1999, showing
characteristic leaf
shapes best
observed pre-anthesis. Column at left: var.
grandiflora,
Allison
11846, from a Bibb County
roadside, typical of the variety in
having
leaf segments only slightly narrowed upward. Center column: var.
harveyana
(Gray)
Sherff,
Allison 11835, from
a glade in Izard County, Arkansas, distinctive in its leaves with
segments
abruptly narrowed above the lower nodes. Right column: var.
inclinata,
Allison 11841, from
the type locality, characterized by, in
addition
to its leaning habit and specialized habitat, a marked tendency to have
fewer leaf segments than
in other varieties.
only slightly narrowed upward. Center column: var.
harveyana
(Gray)
Sherff,
Allison 11835, from
a glade in Izard County, Arkansas, distinctive in its leaves with
segments
abruptly narrowed above the lower nodes. Right column: var.
inclinata,
Allison 11841, from
the type locality, characterized by, in
addition
to its leaning habit and specialized habitat, a marked tendency to have
fewer leaf segments than
in other varieties.
And now we come to
the treatment of Coreopsis in Volume 21 of Flora of North America
North of Mexico (FNA), available for view online at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=108000.
On page 165 of the Castanea article I wrote:
Examination of herbarium specimens was of limited value in understanding the patterns of
variation of outcrop populations of Coreopsis grandiflora.... Had our studies
been limited to the herbarium, we probably would have agreed with Cronquist's taxonomy, and might
have remained uncertain about the distinctiveness of the Ketona Glade populations.
Herbarium specimens of herbaceous plants like Coreopsis grandiflora are virtually always of
flowering or fruiting material. This is entirely understandable, but in rare instances serves to obscure
the differences among taxa, such as those in which the leaf morphology is most distinctive prior to
anthesis. Such is the case with the plants under discussion. Varieties grandiflora,
harveyana, and
inclinata are strikingly different in leaf morphology a few weeks prior to flowering
(see Figure
4),
but the lower leaves are usually withered by the time of anthesis, and so the differences become less
apparent.
On the next page I remarked that
A further
difficulty in interpreting variation in Coreopsis, both in the
herbarium and in the field, is frequent hybridization.... The crossing
experiments of Smith (1976) have shown that varying, often high degrees of
interfertility exist among the various taxa, and it would seem that in many
cases the chief isolating mechanisms are either spatial (allopatry or
differences in habitat preference) or phenological.... The disruption of natural
community boundaries by logging and other land-disturbing activities has
apparently brought into close contact many species of Coreopsis (as
well as Helianthus
L., Silphium, etc.) that were formerly effectively isolated by
differing ecological preferences, and apparent hybrids and hybrid swarms are the
result. In Coreopsis, many of the species seem pre-adapted to
conditions now found along highway and other rights-of-way. Such places often
support populations of plants that seem to combine characters of different taxa.
In the case of Coreopsis grandiflora var. inclinata,
we found putative hybrids with
C. pubescens Ell. at two sites (A. and S. 7633, AUA, UNA,
VDB; A. 11933, JSU, UARK, UNA), one where a road was built across a
glade, the other a glade disturbed in the past by logging.
The taxonomic treatments of the ongoing FNA project are—and
will continue to be—inherently uneven in quality, due to
a number of factors. While the treatments of some genera (e.g., Quercus) are contributed by
recognized experts with extensive relevant experience with the genus, in the
field and in the herbarium, many other treatments (e.g. Carya) are contributed by
non-specialists (at least at the genus level) who must rely on the literature and herbarium specimens. This
situation is unavoidable: given the limited funding available for the project,
it is simply impossible to finance extensive field studies for those plant
groups that require it for their elucidation. This is especially true of genera
like Coreopsis, with dozens of species that moreover have ranges scattered across much of the
continent. While my studies of Coreopsis grandiflora may have resulted in
expertise with that species, I have no comparable understanding of those species
whose ranges lie outside the area where I have decades of field experience,
namely the Southeast. Therefore I harbor no illusion that I could have produced
a better treatment of the genus than that found in FNA.
The treatment of Coreopsis in FNA was contributed by John L.
Strother, Curator of Compositae at the herbaria at the University of
California at Berkeley. This treatment accepted no infraspecific taxa in Coreopsis
grandiflora, which—given the difficulties discussed in my quotations from
the Castanea article—came as no great surprise to me. That it was limited in
approach to literature and herbarium study was hinted at by the lack of any Coreopsis
publications written by its author among the Selected References. A more
explicit indication that the FNA Coreopsis treatment was specimen-based can be found at http://ucjeps.berkeley.edu/people/strother/:
In addition to his
general coordination of treatments as one of three taxon-editors for Compositae,
as a contributor to the flora, Dr. Strother wrote treatments for 119 genera (401
species) and co-wrote treatments of 23 genera (118 species). Most of the work
was specimen-based and was done with collections housed here [UC-Berkeley] in
JEPS and UC. In connection with preparation of treatments for FNANM, Dr.
Strother visited herbaria housed at the Botanical Research Institute of Texas
(BRIT, SMU, and VDB), at the California Academy of Sciences (CAS and DS), and at
the University of Texas (LL and TEX). Dr. Strother is also on the Editorial
Committee for the whole of FNANM and contributes to the editing of all volumes
of the flora.
In addition to the
problems with herbarium collections that l mentioned in the Castanea article, I might have
added that much material of Coreopsis grandiflora (and Silphium)
is seriously biased by the tendency of botanists to collect more often from
roadsides—where hybridization of Coreopsis taxa is
rampant—than from intact natural ecosystems, where it is
rare and the taxa appear more discreet and less likely to exhibit anthropogenic
introgression. As indicated previously, my examination of herbarium specimens at
US and NY in 1993 (and smaller assemblages elsewhere) of C. grandiflora
were more confusing to me than clarifying. In contrast, field observations like
those made of large populations of consistent morphology and referable to var. harveyana
in the spring of 1999 in northern Arkansas, on glades in the Ozarks, were
invaluable to my understanding of infraspecific variation in the species. The
problem with herbarium study of plants such as Coreopsis and Silphium
resulting from lower leaves of distinctive and diagnostic morphology withering by the time of
anthesis is compounded by the tendency of collectors to "top-snatch"
any plant more than a foot or so in height!
seriously biased by the tendency of botanists to collect more often from
roadsides—where hybridization of Coreopsis taxa is
rampant—than from intact natural ecosystems, where it is
rare and the taxa appear more discreet and less likely to exhibit anthropogenic
introgression. As indicated previously, my examination of herbarium specimens at
US and NY in 1993 (and smaller assemblages elsewhere) of C. grandiflora
were more confusing to me than clarifying. In contrast, field observations like
those made of large populations of consistent morphology and referable to var. harveyana
in the spring of 1999 in northern Arkansas, on glades in the Ozarks, were
invaluable to my understanding of infraspecific variation in the species. The
problem with herbarium study of plants such as Coreopsis and Silphium
resulting from lower leaves of distinctive and diagnostic morphology withering by the time of
anthesis is compounded by the tendency of collectors to "top-snatch"
any plant more than a foot or so in height!
Therefore I suppose I should feel grateful that,
instead of being reduced to synonymy without comment, like varieties saxicola
(Alexander) E. B. Smith and harveyana
[photo at right], the Coreopsis grandiflora
discussion in FNA concludes with the statement that "Coreopsis grandiflora var.
inclinata J. R. Allison from glades in Alabama may merit
recognition" (FNA's C. grandiflora treatment is viewable online at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=200023716).
Fortunately for its eventual vindication, var. inclinata is so
edaphically specialized that it does not spread to roadsides and therefore is
far less subject to ongoing "mongrelization" than the other varieties
of C. grandiflora.
As for them (and some of the other well-marked taxa synonymized in FNA without
comment, such as C. pubescens var. robusta), with each passing year the true picture becomes more obscured as
most of the landscape is subjected to ongoing manipulation, with concomitant
introgression. The longer such taxa await systematic studies with a strong field
component, the greater the likelihood that relevant information about their
evolutionary relationships will be lost forever.
Dalea cahaba J. Allison,
named for the Cahaba River, in the tradition
of such Bartram names as Franklinia
alatamaha and Nyssa
ogeche.
All known occurrences of Dalea cahaba are close to the Cahaba
or
tributaries, especially the Little Cahaba River. Click on the
scientific name
to see a close-up of a flowering individual.
 The decumbent
habit of Dalea cahaba (image at left) is
similar to that of the limestone glade endemic,
D. gattingeri
(Heller) Barneby (image at
The decumbent
habit of Dalea cahaba (image at left) is
similar to that of the limestone glade endemic,
D. gattingeri
(Heller) Barneby (image at right). The latter differs markedly
from D. cahaba
by its longer, often sinuous spikes (mostly more
than 2.5 cm long, reaching as much as 7.5 cm), which loosen during and
after anthesis, partially exposing the axis (at least in pressing) and
accompanied by the loss of most of the interfloral bracts. Though its
spikes
are long, its peduncles are short, only 0–3 cm long, while
those
of D.
cahaba are seldom as little as 3
cm long. Furthermore, D.
cahaba
consistently
has 5 leaflets per leaf (rarely 3), while
D. gattingeri often
has
7 or sometimes even 9 leaflets. Dalea
cahaba seems somewhat closer
morphologically to the geographically
more distant D. tenuis (Coult.)
Shinners, a Texas endemic.
right). The latter differs markedly
from D. cahaba
by its longer, often sinuous spikes (mostly more
than 2.5 cm long, reaching as much as 7.5 cm), which loosen during and
after anthesis, partially exposing the axis (at least in pressing) and
accompanied by the loss of most of the interfloral bracts. Though its
spikes
are long, its peduncles are short, only 0–3 cm long, while
those
of D.
cahaba are seldom as little as 3
cm long. Furthermore, D.
cahaba
consistently
has 5 leaflets per leaf (rarely 3), while
D. gattingeri often
has
7 or sometimes even 9 leaflets. Dalea
cahaba seems somewhat closer
morphologically to the geographically
more distant D. tenuis (Coult.)
Shinners, a Texas endemic.
Dalea cahaba shares with D.
tenuis (Coulter) Shinners (image
at right) permanently
dense and conelike spikes mostly less than
2.5 cm long, and
a
similar pattern of bract pubescence. In
addition to
their complete geographic isolation, Dalea
tenuis stands apart
from D. cahaba
by the distinctly retrorse
pubescence of the calyx tube, the almost totally glabrous ovary (pilosulous only
at the style-base), by its long peduncles (as much as 15 cm long,
vs. a
maximum of 8.5 cm in D.
(pilosulous only
at the style-base), by its long peduncles (as much as 15 cm long,
vs. a
maximum of 8.5 cm in D.  cahaba), and its
erect or ascending
habit.
Dalea cahaba,
on the
other hand, has ovaries and pods
that are densely tomentulose on at least the distal two-thirds and a
distinctive
calyx pubescence, the hairs more appressed than in its relatives, and
peculiar
in varying from antrorse to retrorse in orientation on the same calyx,
sinuous
and interwoven (the result a comparatively disheveled calyx vestiture).
Further
differences are found in the calyx teeth, which are often about as
pubescent as
the calyx body in D. cahaba,
usually glabrous or thinly
pilosulous
(though ciliate) in D. tenuis,
and a
decumbent to weakly ascending habit.
cahaba), and its
erect or ascending
habit.
Dalea cahaba,
on the
other hand, has ovaries and pods
that are densely tomentulose on at least the distal two-thirds and a
distinctive
calyx pubescence, the hairs more appressed than in its relatives, and
peculiar
in varying from antrorse to retrorse in orientation on the same calyx,
sinuous
and interwoven (the result a comparatively disheveled calyx vestiture).
Further
differences are found in the calyx teeth, which are often about as
pubescent as
the calyx body in D. cahaba,
usually glabrous or thinly
pilosulous
(though ciliate) in D. tenuis,
and a
decumbent to weakly ascending habit.
The differences among these
species can best be summarized in a key:
Key to Dalea cahaba
and its relatives closest
geographically or morphologically
1
Interfloral bracts with a distinct transverse band of pubescence at the
widest part. Stems mostly 4 dm or more long, branching mostly above the
middle ... Dalea
purpurea.
1
Interfloral bracts with pubescence more generally distributed, never in
a transverse band. Stems less than 4 dm long, branching mostly near or
below the
middle.
2
Calyx
pubescence antrorse. Peduncles less than 3 cm long. Spikes often more
than 2.5 cm long (to 7.5 cm), loosening post-anthesis and losing most
of the interfloral bracts. Leaflets often more than 5 (7 or 9). ... D.
gattingeri.
2
Calyx
pubescence partly or wholly retrorse, especially proximally. Peduncles
mostly more than 3 cm long. Spikes remaining dense, the interfloral
bracts retained between the calyces in fruit. Leaflets nearly always 5.
3
Calyx pubescence
retrorse, the lobes less densely pubescent than
the tube. Ovary and pod glabrous except at the tip. Peduncles often
more
than 8.5 cm long (to 18 cm). Stems mostly erect or
incurved-ascending ... D.
tenuis.
3
Calyx with a
mixture of antrorse and retrorse pubescence, the lobes about as densely
pubescent as the tube. Ovary and pod pubescent over a majority of the
surface. Peduncles at most 8.5 cm long. Stems decumbent to weakly
ascending ... D.
cahaba.
Erigeron strigosus Muhl. ex Willd. var. dolomiticola J. Allison and
var. calcicola
J. Allison. The Latin suffix -icola
means
"-dweller" or "-inhabitant," and the names mean "dolomite dweller" and
"limestone dweller,"
respectively. The ending of these epithets in -a,
rather than -us
reflects the fact that they are nouns, not adjectives. Click on "var. dolomiticola,"
above, to see
an image of a flowering individual with a basal
rosette-cluster that is admittedly more strongly developed than is
usual (the
image of var. dolomiticola
in the second paragraph below is
closer to
average in this respect). Clicking on "var. calcicola,"
above,
shows
a plant at the latter's type locality in Rutherford County, Tennessee
(with Ruellia
humilis Nutt.).
 The
Ketona Glade endemic seemed clearly aligned with Erigeron
strigosus
and E. annuus
(L.) Pers. in having disk
flowers with a double
pappus, with an
The
Ketona Glade endemic seemed clearly aligned with Erigeron
strigosus
and E. annuus
(L.) Pers. in having disk
flowers with a double
pappus, with an outer
series of setose scales and inner series of capillary bristles, and
with ray
flowers
bearing only the
scales (see image at
right). The tufts of remarkably narrow radical leaves seemed
distinctive, approached in slenderness only by the very
narrowest-leaved
extremes of E. strigosus Muhl.
ex Willd. var. beyrichii (Fisch.
& C. A. Mey.) Torr. & Gray ex Gray, as found
in xeric
habitats such as
sand ridges. Variety beyrichii
is, however, like all
varieties of "daisy fleabane"
described before 2001, an annual
or sometimes a biennial. The plant
of var. beyrichii shown
at left is typical in that the
leaves of the basal rosette have withered
by the time anthesis is well under way (detail, right side of image).
outer
series of setose scales and inner series of capillary bristles, and
with ray
flowers
bearing only the
scales (see image at
right). The tufts of remarkably narrow radical leaves seemed
distinctive, approached in slenderness only by the very
narrowest-leaved
extremes of E. strigosus Muhl.
ex Willd. var. beyrichii (Fisch.
& C. A. Mey.) Torr. & Gray ex Gray, as found
in xeric
habitats such as
sand ridges. Variety beyrichii
is, however, like all
varieties of "daisy fleabane"
described before 2001, an annual
or sometimes a biennial. The plant
of var. beyrichii shown
at left is typical in that the
leaves of the basal rosette have withered
by the time anthesis is well under way (detail, right side of image).
It was apparent
right
away when
I first
saw it in flower, in June 1992, that the race of Erigeron
strigosus
that was quite frequent on  the Ketona Glades was different from the
common,
weedy daisy fleabanes that are familiar wildflowers (weeds to some!)
of roadsides. The glade plants
the Ketona Glades was different from the
common,
weedy daisy fleabanes that are familiar wildflowers (weeds to some!)
of roadsides. The glade plants had basal offsets (see image at left) of
very narrow, erect leaves that were fresh and green—anything
but senescent—at
anthesis and, moreover, occasional dead remains of stems of the
previous
year were also associated with the flowering stems. It seemed that
these
glade plants must be perennial(!), especially as visits in the summer
and
fall showed that the rosettes remained green after the flowering stems
of the early- and mid-season senesced. Once freezing weather arrived,
the
rosettes remained green but became laxer and more prostrate.
had basal offsets (see image at left) of
very narrow, erect leaves that were fresh and green—anything
but senescent—at
anthesis and, moreover, occasional dead remains of stems of the
previous
year were also associated with the flowering stems. It seemed that
these
glade plants must be perennial(!), especially as visits in the summer
and
fall showed that the rosettes remained green after the flowering stems
of the early- and mid-season senesced. Once freezing weather arrived,
the
rosettes remained green but became laxer and more prostrate.
But before
winter had
come to
Bibb
County I made a trip to Nashville to visit the herbarium at Vanderbilt
University, to examine the collection there, rich in specimens from
Alabama,
and to discuss taxonomic
matters with its curator (and collector of the
majority
of its specimens), Dr. Robert Kral. Naturally
I also took advantage of the opportunity to visit some of the limestone
"cedar" glades so well developed in that part of Tennessee (it was
familiarity
with these, from visits beginning early in the 1980s, that helped me to
recognize, when the time came, that many of the dominant Ketona Glade
plants
weren't the usual denizens of calcareous glades). During my previous
explorations
of limestone glades I'd never paid much attention to Erigeron
strigosus—why
bother with a "weed" when there were so many curious endemics to enjoy?
But,
having found a new, perennial variety on dolomite glades in Bibb
County,
Alabama, I naturally paid a little more attention to daisy fleabanes
when
I revisited the
Middle Tennessee cedar  glades
on October 27, 1992. To my
surprise, I found that one of the most typical plants of sunny places
on
these glades was yet another perennial
form of E, strigosus.
Despite the fact that there were lingering patches of ice from a frost
of the preceding night, the senescing but still sparingly floriferous
stems
of E. strigosus
were associated with fresh, green, obviously
overwintering
rosettes (center of image at right—the color version of
glades
on October 27, 1992. To my
surprise, I found that one of the most typical plants of sunny places
on
these glades was yet another perennial
form of E, strigosus.
Despite the fact that there were lingering patches of ice from a frost
of the preceding night, the senescing but still sparingly floriferous
stems
of E. strigosus
were associated with fresh, green, obviously
overwintering
rosettes (center of image at right—the color version of Figure 7
from the Castanea
paper—with rosettes indicated by
arrows). As with
the Ketona Glade variety, careful extraction of the plants showed that
the offsets were connected to the flowering stems (as in the upper left
portion
of the image, showing the lower part of a pressed specimen). While var. dolomiticola
had the additional distinction of extremely narrow basal leaves (upper
right portion of image), these cedar glade perennials had basal leaves
that were closer in shape to those of the common, weedy, annual kinds.
I asked myself, "Could it be that the glade habitat was somehow
responsible,
rather than a genetic difference, for the perennial duration of these
plants?"
But no, once I noticed that the cedar glade perennial was further
distinguished
by being consistently much less pubescent than other Erigeron
strigosus
varieties—the
cauline leaves glabrous except for marginal cilia and sparse,
strigillose
hairs along the midvein—it was clear that I had found a
distinctive and
unappreciated cedar glade endemic.
Figure 7
from the Castanea
paper—with rosettes indicated by
arrows). As with
the Ketona Glade variety, careful extraction of the plants showed that
the offsets were connected to the flowering stems (as in the upper left
portion
of the image, showing the lower part of a pressed specimen). While var. dolomiticola
had the additional distinction of extremely narrow basal leaves (upper
right portion of image), these cedar glade perennials had basal leaves
that were closer in shape to those of the common, weedy, annual kinds.
I asked myself, "Could it be that the glade habitat was somehow
responsible,
rather than a genetic difference, for the perennial duration of these
plants?"
But no, once I noticed that the cedar glade perennial was further
distinguished
by being consistently much less pubescent than other Erigeron
strigosus
varieties—the
cauline leaves glabrous except for marginal cilia and sparse,
strigillose
hairs along the midvein—it was clear that I had found a
distinctive and
unappreciated cedar glade endemic.
Subsequent
visits to
limestone
glades in northern
Alabama and to a single glade in Floyd County, Georgia (a Georgia plant
shown
at right) revealed that the perennial Erigeron
strigosus of
limestone
glades was by no means restricted to Middle
Tennessee. Oddly, the plant was not seen on any of the cedar glades
in Catoosa County, Georgia, where the characteristic flora is best
developed
in my home state.
In November of 2001, I shipped to Richard D. Noyes (University of
Colorado) live material I collected of Erigeron strigosus varieties dolomiticola
(two sites) and calcicola (from two sites each in Alabama and
Tennessee) for use in evolutionary studies of Erigeron sect. Phalacroloma.
These provided helpful data for at least two publications to date, Noyes &
Allison 2005, "Cytology, Ovule Development, and Pollen Quality in Sexual Erigeron
strigosus (Asteraceae)" [International Journal of Plant Science 166(1):
49–59 (available at http://faculty.uca.edu/rnoyes/PDFs/IJPS.2005.pdf)], and Noyes 2006, "Intraspecific Nuclear Ribosomal DNA
Divergence and Reticulation in Sexual Diploid Erigeron strigosus"
[American Journal of Botany 93(3): 470–479 (available at http://faculty.uca.edu/rnoyes/PDFs/AJB.2006.pdf)]. Among other things,
our 2005
paper reported that both varieties were diploid (2n=18) and sexual (as was a
third, unnamed variety). Noyes' 2006 paper presented evidence that "the three
groups of sexual plants form separate monophyletic clades and that edaphic
specialization is ancestral in the group." Noyes stated that:
Sexuality
in E. strigosus var. calcicola and E. strigosus var. dolomiticola
is particularly noteworthy. Because these taxa are now confirmed to be diploid
and sexual rather than polyploid and apomictic they conform to more widely
accepted units of biodiversity. As such, these taxa unquestionably can be added
to the substantial list of plant taxa endemic to glade habitats in the
southeastern United States (Estill and Cruzan 2001,* Baskin and Baskin 2003**).
*
Estill, J.C. and M.B. Cruzan. 2001. Phytogeography of rare plant species endemic
to the southeastern United States. Castanea 66: 3–23.
**
Baskin, J.M. and C.C. Baskin. 2003. The vascular flora of cedar glades of the
southeastern United States and its phytogeographical relationships. J Torrey Bot
Soc 130:101–118.
Another interesting observation of Noyes' 2006 paper was that
The rDNA phylogeny
for sexual diploid Erigeron strigosus (Figs. 2, 3) provides insight into
the evolution of sexual populations of Erigeron sect. Phalacroloma.
First, E. strigosus var. calcicola occurs basal to E. strigosus
var. dolomiticola and sexual E. strigosus var. strigosus.
This is consistent with the hypothesis that the glade endemics, which are
perennial through the production of stolons or overwintering rosettes, are
ancestral to the typical annual or weakly perennial forms of E. strigosus,
including sexual E. strigosus var. strigosus (Allison and Stevens,
2001). Glades in the southeastern United States are harsh and variable
environments, prone particularly to extreme heat and desiccation (Baskin and
Baskin, 1999*). The perennial habit may foster survival and reproduction during
prolonged water stress. This phylogenetic pattern indicates that the
nonspecialized habit of sexual E. strigosus var. strigosus is
likely derived from an ancestor that was an edaphic specialist. This is counter
to the conventional model that posits that edaphic plant specialists usually
evolve from nonspecialist progenitors (MacNair and Gardner, 1998;** Rajakaruna,
2004***).
*
Baskin, J. M., AND C. C. Baskin. 1999. Cedar glades of the southeastern United
States. In R. C. Anderson, J. S. Fralish, and J. M. Baskin [eds.], Savannas,
barrens, and rock outcrop plant communities of North America, 206–219.
Cambridge University Press, Cambridge, UK.
**
MacNair, M. R. and M. Gardner. 1998. The evolution of edaphic endemics. In D. J.
Howard and S. H. Berlocher [eds.], Endless forms: species and speciation, 157–171.
Oxford University Press, New York, New York.]
***
Rajakaruna, N. 2004. The edaphic factor in the origin of plant species.
International Geology Review 46: 471–478.
The treatment of Erigeron
in FNA (Vol. 20, 2006) was contributed by Guy L. Nesom of the Botanical Research Institute of
Texas. Nesom's FNA treatment of Erigeron strigosus recognized the
two perennial outcrop endemic varieties but var. beyrichii (which
was characterized on p. 173 of the Castanea article as "weakly
differentiated) was reduced to synonymy under var. strigosus. The FNA
treatment of E. strigosus var.
dolomiticola treatment is available online at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=250068373,
that of var. calcicola at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=250068372.
| Castanea
Supplement, Part 2 of 2 |

|
| Home |
Castanea
paper Table of Contents |
 |
 |
|
Most recent update: January 12, 2010
 Castilleja
coccinea
Castilleja
coccinea


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}