Supplement to a paper published in Castanea, Vol. 66(1-2):154–205,
March/June 2001
Musical selection: Gavotte et Doubles from Nouvelles Suites
de Pièces de Clavecin by Jean-Philippe Rameau (1683-1764)
Vascular Flora
of Ketona Dolomite Outcrops in Bibb County, Alabama
(CONTINUED)
Liatris oligocephala J. Allison,
from the Latinized Greek oligo-,
"few" and
-cephala,
"-headed;" so named because the majority of plants in the wild bear
only
1 to 3 heads, as shown in the image below, right. Click on the
scientific name,
above, to see an image of a group of vigorous flowering individuals,
most of them with more than the
average number of heads.
 Liatris oligocephala
is the only known Liatris lacking
tiny bristles on the ribs of its achenes. It is also the only Liatris
that
produces mostly one to
Liatris oligocephala
is the only known Liatris lacking
tiny bristles on the ribs of its achenes. It is also the only Liatris
that
produces mostly one to three heads or at most a simple cyme. Liatris
cymosa
(Ness.) K. Schum. and L. ohlingerae
(Blake) B. Rob. often
have the
heads arranged in a simple cyme, but vigorous individuals of both these
species
bear compound cymes. Despite its unique features, the Ketona Glade
plant is
clearly a Liatris
because of its stocky, often globose "corm,"
lack of any bracts between the florets, and the fact that it hybridizes
with L. cylindracea.
three heads or at most a simple cyme. Liatris
cymosa
(Ness.) K. Schum. and L. ohlingerae
(Blake) B. Rob. often
have the
heads arranged in a simple cyme, but vigorous individuals of both these
species
bear compound cymes. Despite its unique features, the Ketona Glade
plant is
clearly a Liatris
because of its stocky, often globose "corm,"
lack of any bracts between the florets, and the fact that it hybridizes
with L. cylindracea.
At right is the color version of Figure 9 from the Castanea
paper.
It
relates to hybridization in Ketona Glade species of Liatris.
Top
row,
photographs taken 13 July 1993 at the type locality of
L. Xfreemaniana
(L.
cylindracea X L.
oligocephala),
showing differences in head arrangement: A.
L.
cylindracea. B. L. Xfreemaniana.
C.
L. oligocephala.
Middle row, pressed heads of same taxa, from
collections [UNA] made 18 July 1993, showing differences in outer
phyllary
shape and in degree of ciliation: D.
Allison & Stevens 7800.
E. Allison & Stevens
7801. F. Allison
& Stevens 7815.
Bottom row, photographs taken 19 August 1999 at the type locality of
L. Xmacdanieliana
(L. cylindracea X L.
squarrosa), showing differences
in phyllary shape and orientation:
G.
L. cylindracea.
H. L. Xmacdanieliana.
I.
L. squarrosa.
A 2005 paper by T. M. ("Mike') Hardig, James R. Allison and Edward E. Schilling
was focused on Liatris oligocephala, L. cylindracea and
putative hybrids (L. Xfreemaniana). Titled "Molecular
Evidence of Hybridization between Liatris oligocephala (Asteraceae) and
More-Widespread Congener: A Preliminary Assessment of the Potential for
Extinction" (Castanea 70:246-254, available at http://www.bioone.org/doi/pdf/10.2179/0008-7475%282005%29070%5B0246%3AMEOHBL%5D2.0.CO%3B2),
its abstract is quoted here in entirety:
Hybridization may
increase the risk of extinction for some rare and endangered plant species,
either by increasing the rate of random genetic drift affecting populations or
by introgression-mediated genetic assimilation. The former may occur when
hybrids are less fit than non-hybrids and backcrossing would be less likely to
occur, the latter may occur when hybrids are as fit, or fitter, than parents,
and backcrossing is fairly common. To assess the risk of extinction by
hybridization one must assess whether hybridization is actually occurring, and,
if so, whether the hybrids are sterile, or are backcrossing. We analyzed one
population each of the rare Liatris oligocephala and L. cylindracea
and seven field-identified hybrids for ribosomal DNA and chloroplast DNA
markers, and morphological characteristics to make a preliminary determination
of the potential for L. oligocephala extinction via hybridization. We
found unequivocal evidence that the two species are hybridizing, but no
unequivocal evidence of backcrossing.
Further study will be necessary to determine whether Liatris oligocephala
is threatened at all by genetic "swamping," which could result from
backcrossing from F1 hybrids with L. cylindracea. Greater impact to L.
oligocephala populations may well come from competition with F1 hybrids (L. Xfreemaniana)
in microsites that are transitional from the severe microhabitats to which L.
oligocephala seems better adapted to those microhabitats more favorable to L.
cylindracea, with deeper soil and/or more shade.
That same year (2005), Guy L. Nesom published a paper titled
"Infrageneric
classification of Liatris (Asteraceae: Eupatorieae)" [Sida 21:1305-1321
(available for download at http://www.guynesom.com/LiatrisInfragClass.pdf)].
In the Castanea paper (p. 178) I had written that "This new Liatris
appears to be sufficiently distinctive to merit the establishment of a new
series, perhaps even a new section, to accommodate it, but we will leave
that decision to a future monographer." In his 2005 paper, Nesom made L.
oligocephala the type-species of his newly created
Section Vorago. I
was surprised to find a second species included there with it, the Texas
endemic L. tenuis Shinners, a plant quite dissimilar to L.
oligocephala in gross
appearance. I suspect that future molecular studies will not support this
disposition; this would apparently come as no surprise to Nesom, either, as
he was explicit that the association of these two species was tentative:
a new
series, perhaps even a new section, to accommodate it, but we will leave
that decision to a future monographer." In his 2005 paper, Nesom made L.
oligocephala the type-species of his newly created
Section Vorago. I
was surprised to find a second species included there with it, the Texas
endemic L. tenuis Shinners, a plant quite dissimilar to L.
oligocephala in gross
appearance. I suspect that future molecular studies will not support this
disposition; this would apparently come as no surprise to Nesom, either, as
he was explicit that the association of these two species was tentative:
Liatris
oligocephala is similar to L. tenuis in features of the involucre,
corolla, and pappus, but because no synapomorphy is evident, it seems likely
that these two species originated independently from the ancestral stock of
sect. Liatris. Thus, sect. Vorago, as delimited here, may not be
monophyletic.... The name of the section (“vorago,” Latin, gulf) alludes to
the range of the two species on the Gulf Coastal Plain as well as to the
considerable morphological “gulf” between them."
Nesom's choice of the epithet Vorago was rather unfortunate, as the
type species, Liatris oligocephala is not found on the Gulf
Coastal Plain, but instead in the Ridge and Valley Province (its obligate
association with a Cambrian-age sedimentary rock formation makes this
unequivocal). Nesom may have gotten this misconception from certain literature
containing the same error, perhaps owing to the fact that the majority of Bibb
County, Alabama does in fact lie within the Gulf Coastal Plain.
Guy L Nesom also contributed the FNA treatment of Liatris (Vol. 21,
2006). His treatment of L. oligocephala is available online at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=250067110.
Unlike full species, hybrids are of course not treated individually. Those that
have received names as nothospecies, including L. Xfreemaniana and
L. Xmacdanieliana, are listed in FNA with their putative (sometime
speculative) parents (as can be viewed at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=242416766).

Onosmodium decipiens J. Allison.
The Latin word decipiens
means
"deceiving;"
so named because the species exhibits features alternatively of either
O.
virginianum (L.) A. DC. or of O.
molle Michx. ssp. hispidissimum
(Mackenzie) Boivin, which caused early collections to be labeled
as one or the other of those taxa. Click on the scientific name to see
an image
of a flowering individual.
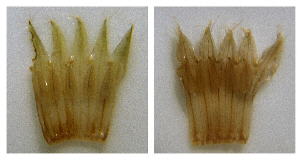 In stature and
flower
color, Onosmodium
decipiens more closely resembles
O. virginianum.
The former
has longer hairs on its stem (> 2.5 mm vs. <2
In stature and
flower
color, Onosmodium
decipiens more closely resembles
O. virginianum.
The former
has longer hairs on its stem (> 2.5 mm vs. <2 mm), but
the
critical
differences are
in the flowers. The
photo at left shows
dissected corollas rehydrated
from
pressed specimens of O.
virginianum (left half) and O.
decipiens
(right half). The consistently shorter filaments of O.
virginianum
cause the anther apices to be positioned distinctly below the sinuses
(notches) of
the corolla, while the consistently longer filaments of O.
decipiens
result in anther apices that reach—and often slightly
exceed—the bases
of the sinuses. A further difference is seen in the shape of the
corolla
lobes, more narrowly and sharply triangular in O.
virginianum
than
in O. decipiens.
mm), but
the
critical
differences are
in the flowers. The
photo at left shows
dissected corollas rehydrated
from
pressed specimens of O.
virginianum (left half) and O.
decipiens
(right half). The consistently shorter filaments of O.
virginianum
cause the anther apices to be positioned distinctly below the sinuses
(notches) of
the corolla, while the consistently longer filaments of O.
decipiens
result in anther apices that reach—and often slightly
exceed—the bases
of the sinuses. A further difference is seen in the shape of the
corolla
lobes, more narrowly and sharply triangular in O.
virginianum
than
in O. decipiens.
The corollas of Onosmodium
decipiens
are more like those of O. molle ssp. hispidissimum,
except that they are pale yellow (subspecies of O.
molle have
white
corollas with greenish-tinged  lobes). The pictures above, right (O.
decipiens)
and at left (O. molle ssp. hispidissimum)
show
plants early
in anthesis. The marked differences
between these two species in habit (ascending vs. erect) and in average
stature (most plants less than 6.5 dm
lobes). The pictures above, right (O.
decipiens)
and at left (O. molle ssp. hispidissimum)
show
plants early
in anthesis. The marked differences
between these two species in habit (ascending vs. erect) and in average
stature (most plants less than 6.5 dm in O.decipiens,
with a maximum of 8.4 dm, while plants a meter or more tall are
frequent in subspecies of O.
molle) are apparent. Also
evident is the tendency for O.
decipiens (like O.
virginianum), to produce several
stems, while taxon hispidissimum
(like other subspecies of O.
molle), produces one or very
few.
While the long, stiff
pubescence of both taxa appears similar, a look at the foliage with a
hand
lens reveals another distinction. Between the veins, taxon
hispidissimum
has an under-pubescence of shorter, more appressed hairs beneath the
long,
spreading, conspicuous ones
(another common feature of the subspecies of O.
molle), while
in O. decipiens
(and O. virginianum)
such hairs
are confined to the veins. A
further difference between these two is found in the nutlets, which are
smaller and gradually tapered to the truncate base in O.
decipiens
(left half of image at right, above), while nutlets with distinctly
"collared" bases have always been
considered a hallmark of taxon hispidissimum
(right half of
image at
right, above).
in O.decipiens,
with a maximum of 8.4 dm, while plants a meter or more tall are
frequent in subspecies of O.
molle) are apparent. Also
evident is the tendency for O.
decipiens (like O.
virginianum), to produce several
stems, while taxon hispidissimum
(like other subspecies of O.
molle), produces one or very
few.
While the long, stiff
pubescence of both taxa appears similar, a look at the foliage with a
hand
lens reveals another distinction. Between the veins, taxon
hispidissimum
has an under-pubescence of shorter, more appressed hairs beneath the
long,
spreading, conspicuous ones
(another common feature of the subspecies of O.
molle), while
in O. decipiens
(and O. virginianum)
such hairs
are confined to the veins. A
further difference between these two is found in the nutlets, which are
smaller and gradually tapered to the truncate base in O.
decipiens
(left half of image at right, above), while nutlets with distinctly
"collared" bases have always been
considered a hallmark of taxon hispidissimum
(right half of
image at
right, above).
The major differences among
these taxa of Onosmodium
are
summarized in
the following table:
|
Morphological
comparison
of three Onosmodium
taxa
|
O.
molle ssp. hispidissimum
|
O. decipiens
|
O. virginianum
|
|
stem
length
|
to ca. 12 dm
|
to ca. 6.5 (-8.4) dm
|
to ca. 5 dm
|
|
stems
per plant
|
few
|
few to several
|
few to
several |
|
stem
hair length
|
> 2.5 mm
|
> 2.5 mm
|
< 2.0 mm
|
|
leaf
indument
|
double
|
simple
|
simple
|
|
corolla
color
|
white w/ greenish lobes
|
light yellow
|
light yellow
|
|
corolla
lobes
|
nearly deltoid
|
nearly deltoid
|
acuminate
|
|
anther
apices
|
at corolla sinuses
|
at sinuses
|
below sinuses
|
| nutlet
size |
large
|
small
|
small
|
|
nutlet
base
|
constricted
|
unconstricted
|
unconstricted
|
The tenth annual (2008) Spring Symposium at Eastern Oregon
University (La Grande) included a poster presentation by Kali Miller,
titled "Comparing Pyrrolizidine Alkaloids in Onosmodium decipiens
and O. virginianum." I am not aware of any publication of results of
this study. The abstract (found at www.eou.edu/sprsymp/documents/program.pdf)
is as follows:
Pyrrolizidine
alkaloids (PA’s), potentially toxic natural secondary products, are typically
found in the plant family Boraginaceae. As a member of the family, the genus Onosmodium
can be expected to contain PA’s. Onosmodium virginianum is found
throughout the southeastern United States, while the recently described closely
related O. decipiens is prevalent only in Bibb County, Alabama. Previous
work on O. decipiens established the presence of the PA’s uplandicine
and lithospermine. A preliminary GC/MS investigation of O. virginianum
has discovered the main PA to be a novel saturated analog of the PA echimidine
with lesser amounts of the PA’s uplandicine, intermidine, echimidine, and
supinine. The exact structure of the alkaloids found in the O. virginianum
will be elucidated using NMR experiments. A chemosystematics comparison between
the two Onosmodium species will be presented.
Silphium
glutinosum J. Allison
and S.
perplexum J. Allison,
named with Latin
adjectives meaning "sticky" and "confused,"
respectively. The copious, glandular indument
of S. glutinosum
(and of S. perplexum)
gives the plants
a sticky feel. After removing specimens from the plant press, not only
are the newspapers they were pressed between usually stained yellow but
the plants stick to the paper. Fortunately, they are easily peeled away
from the paper, producing a sound not unlike that made by separating
Velcro®.
Naming the latter, Black
Belt species
"confused" seemed apt for a plant that appeared unable to make up its
mind
whether it wanted to be S.
glutinosum or S.
integrifolium
Michx. (then, too, early collections—dating back as far as
1885—were confused
with the last-named
species). Click on the scientific names, above, for images of plants in
flower.
 The combination of gradually
reduced leaves, and heads usually with
13 rays align Silphium
glutinosum with the S. asteriscus
complex (e.g.,
The combination of gradually
reduced leaves, and heads usually with
13 rays align Silphium
glutinosum with the S. asteriscus
complex (e.g., S. asteriscus
L., S. trifoliatum
L. and their varieties), but none of these
has
glandular stems, leaves, or involucres, and their leaves average
narrower than
those of S. glutinosum.
S. asteriscus
L., S. trifoliatum
L. and their varieties), but none of these
has
glandular stems, leaves, or involucres, and their leaves average
narrower than
those of S. glutinosum.
After showing Robert Kral Silphium
glutinosum in the field
in Bibb
County, prior to anthesis, he called attention to densely glandular
specimens labeled S.
integrifolium he had collected
from the Black Belt province of
Alabama, in
Dallas County (Kral 48820,
VDB) and Perry County (Kral
47891,
VDB). Tim Stevens and I visited both of these populations, and
"windshield
surveying" along highways and back roads has resulted to date in the
discovery of well
over a dozen additional populations of a densely glandular Silphium
in
those two counties. After comparison of living populations and
preserved
material of these plants with S. integrifolium
and with S.
glutinosum, we determined that a
suite of characters exists that
distinguish
the glandular Silphium of
Dallas and Perry Counties from
either of those species.
Silphium
integrifolium differs from S.
glutinosum in
many
respects. The former is, under favorable conditions in the wild and in
the
garden (e.g., garden of Allison), a taller plant with nodes slightly
closer
together and therefore more numerous. Those leaves that persist until
anthesis
in S. integrifolium are
normally sessile and, like the stem,
eglandular.
It also has a narrower inflorescence, on average with fewer but larger
heads,
these with consistently more numerous rays and phyllaries (the latter
never
with the revolute margins seen in S.
glutinosum) and with acute
receptacular bracts (pales or chaff).
Plants of the Dallas and Perry
County Silphium
are
intermediate
between S. glutinosum and
S. integrifolium
for some
characters and
for others resemble one or the other of those species, as indicated in
the
following table:
|
Morphological
comparison of
three Silphium
taxa
|
S.
glutinosum
|
S.
perplexum
|
S.
integrifolium
|
|
plant
height
|
mostly 8-15 dm
|
mostly 15-20 dm
|
often >15 dm
|
|
nodes
below inflorescence
|
mostly 6 or 7
|
mostly 9-12
|
often 10 or more
|
|
stem
and leaf glandular hairs
|
long
|
short
|
absent
|
|
lowest
petiole length at
anthesis
|
mostly 6-15 cm
|
mostly 5-8 cm
|
mostly 0 cm
|
|
inflorescence
|
open
|
open or ±
congested
|
±
congested
|
|
chaff
apices
|
obtuse
|
obtuse to acutish
|
strongly acute
|
|
number
of rays per head
|
(8) 12-14 (16)
|
(17) 19-23 (33)
|
(16) 21-34 (35)
|

At left is the color version of
Figure
12 from the Castanea
paper. Top row, Silphium
glutinosum,
Bibb County, Alabama: A. Habit,
including open inflorescence; "Desmond's Glade," 9 September 1993. B.
Head,
frontal view, with the normal 13 rays; "Goat Glade South," 14 June
1992.
C. Head, lateral view, showing outer phyllaries the longest and with
recurving
margins; "Browne's Dam North Glade West," 3 July 1992. Middle row, S.
perplexum: D. Habit; Perry
County, Alabama, Co. Rd. 6, 19 August
1999.
E. Heads, frontal view, showing maximal no. of rays per head; same date
and location as in D. F. Head, three-quarters view showing the species'
comparatively "ordinary" involucre; Dallas County, Alabama, type
locality,
18 August 1999. Bottom row, S.
integrifolium: G. Habit,
including
more numerous nodes, Lowndes County, Mississippi, Alt. U.S. Hwy. 45, 9
August 1992. H. Heads, frontal view, with numerous rays; Sumter County,
Alabama, Ala. Hwy. 17, 19 August 1999. I. Comparatively congested
inflorescence,
same date and location as in G.

The
image
at lower left shows the chaff (pales,
receptacular bracts) from a
rehydrated and dissected
head of Silphium  glutinosum.
The chaff apices are consistently
obtuse.
The chaff of S. integrifolium
(right) is dramatically
different—strongly
acute. While intermediate between these two species in many respects, S.
perplexum tends to have chaff
apices more similar to those of S.
glutinosum.
glutinosum.
The chaff apices are consistently
obtuse.
The chaff of S. integrifolium
(right) is dramatically
different—strongly
acute. While intermediate between these two species in many respects, S.
perplexum tends to have chaff
apices more similar to those of S.
glutinosum.
The treatment of Silphium in
FNA (Vol. 21, 2006) was contributed by Jennifer A. Clevinger,
of James Madison University. Her treatment of S. glutinosum is
available online at http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=250067527.
As stated on page 187 of the Castanea article, "Given the frequency of
hybridization in human-perturbed habitats, it appears that, among
related taxa of Silphium, barriers
to gene flow are chiefly ecological. Alteration of the landscape by
humans appears to have disrupted
these barriers, just as in Coreopsis." It follows that any
taxonomic treatment of Silphium faces similar impediments to a thorough
understanding to those confounding a treatment of Coreopsis, as discussed
above under that genus. Except that the mostly tall Silphium spp. are
even more subject to be represented by "top-snatched" material in
herbaria! The mostly herbarium-based Silphium treatment in FNA reflects this
unfortunate state of affairs. In particular the elements of the S. asteriscus
complex, perhaps necessarily "lumped" in large part as a practicality,
are desperately in need of studies that include a strong field component that
focuses on habitats not created or transformed by humans, a quest that becomes
increasingly difficult, if not impossible, every day.
I would feel better about the failure to accept Silphium
perplexum in FNA if what was written about it there were more accurate.
According to Clevinger, "James Allison (in protologue) suggested
that Silphium perplexum is a hybrid between S. glutinosum and S.
integrifolium. It has glandular hairs on its stems, leaves, and phyllaries
as in S. glutinosum; it is also found on chalk, has taller stems, larger
leaf blades, and deeper yellow corollas. Further study is needed to determine
its status." Most unfortunately, the most salient difference between S.
glutinosum and S. perplexum received no mention, namely the
consistently greater number of ray flowers in the latter (or ovaries, as can be
determined by dissection of a head once some of the ray flowers have abscised).
It is unusual for the number of rays/ovaries in a head of S. glutinosum
to number other than 13, unheard of for S. perplexum to have so few. I
interpreted this and other features of their comparative morphology as well as
their geographic ranges as suggestive of an origin of S. perplexum
in prehistoric hybridization of S. glutinosum with the [today]
wholly allopatric S. integrifolium. This is not at all the same thing as
calling Silphium perplexum "a hybrid between S. glutinosum and S.
integrifolium" as misstated by Clevinger, which would imply that these
two allopatric species are in genetic contact today or even in the recent
past. All three species are in fact allopatric.
stems, leaves, and phyllaries
as in S. glutinosum; it is also found on chalk, has taller stems, larger
leaf blades, and deeper yellow corollas. Further study is needed to determine
its status." Most unfortunately, the most salient difference between S.
glutinosum and S. perplexum received no mention, namely the
consistently greater number of ray flowers in the latter (or ovaries, as can be
determined by dissection of a head once some of the ray flowers have abscised).
It is unusual for the number of rays/ovaries in a head of S. glutinosum
to number other than 13, unheard of for S. perplexum to have so few. I
interpreted this and other features of their comparative morphology as well as
their geographic ranges as suggestive of an origin of S. perplexum
in prehistoric hybridization of S. glutinosum with the [today]
wholly allopatric S. integrifolium. This is not at all the same thing as
calling Silphium perplexum "a hybrid between S. glutinosum and S.
integrifolium" as misstated by Clevinger, which would imply that these
two allopatric species are in genetic contact today or even in the recent
past. All three species are in fact allopatric.
I am much less concerned about the eventual vindication of Silphium
perplexum as distinctive as I am about the edaphically less specialized taxa
regarded as varieties of S. asteriscus. While introgression of S.
perplexum with other species is indeed occurring along roadsides—a
hybrid swarm within the city limits of Selma (west side of Bell Rd, ca. 0.2 rd.
mi. S of jct. U.S. Hwy. 80) is especially memorable)—a
sizeable amount of its chalk prairie habitat around the type locality of Old
Cahaba (Dallas County) is now protected (another
3,020 acres as recently as September 2003), and hybrids are unlikely to have
greater fitness there than S. perplexum, except along roadsides.
That Silphium perplexum is as about as close
morphologically to S. integrifolium as it is to S. glutinosum (and
thus can satisfactorily be made a mere variety of neither) is borne out by
Clevinger's 1999 annotation of the only 19th century collection known of S.
perplexum (at GH, shown at right) as S. integrifolium (likewise
determined by
all other annotators).
An additional factual error in the FNA treatment of Silphium
begs
correcting: the habitat of S. glutinosum is given there as "Limestone
glades, along rocky streams." While a very few occurrences along rocky
streams are over limestone (e.g., near Pratt's Ferry, as mentioned above,
immediately preceding the section titled "Biological Communities), the
glades on which S. glutinosum grows are always underlain by dolomite. While
limestone and dolomite are both calcareous sedimentary rocks, considering the
terms interchangeable (as also in Lawless et al., 2006) seems analogous to
regarding the words "daisies" and "goldenrod" equivalent
because they belong to the same botanical family!

In discussing Spigelia gentianoides Chapm. in A. DC., the
Castanea paper indicated that
Many, if not
most, Florida populations of Spigelia gentianoides
have been lost or are in the process of disappearing as a result of fire
suppression. By contrast, the Ketona Glade habitat is probably maintained more
by rockiness and edaphic conditions. However, occasional wildfire in the
ecotones would have served to limit encroachment from the glade margins.
Such a role in the past for wildfire in maintaining the Ketona Glade ecosystem
is indicated by the persistence, in the forest matrix surrounding some of the
glades, of longleaf pine and such wiregrass-country associates as Sericocarpus
tortifolius (Michx.) Nees and, less commonly, Carphephorus
odoratissimus (J. F. Gmel.) Herbert.
A 2007 paper examined
the effects of prescribed fire on rare plants of Ketona dolomite glade
communities. Titled "The Effect of Fire Reintroduction on Endemic and Rare
Plants of a Southeastern Glade Ecosystem" (Restoration Ecology16:39-49),
this paper by R. Scot Duncan, Corinna B. Anderson, Heather N.
Sellers, and Erin E. Robbins, is available from http://www3.interscience.wiley.com/cgi-bin/fulltext/119413438/PDFSTART.
The abstract is quoted
in full as follows:
Open habitats
dominated by herbaceous plants on thin, rocky soils occur within the forests of
eastern North America. Although these habitats vary in origin, structure,
geology, and species composition, all contribute greatly to regional
biodiversity by harboring endemic and/or rare plants. Little is known about how
disturbances affect plant populations in these ecosystems. Fire once was a
frequent natural disturbance in the Ketona dolomite glades of Alabama, an
ecosystem harboring eight endemic taxa and numerous other species of
conservation concern. We designed an experiment to determine how the
reintroduction of fire into the glades and surrounding longleaf pine forests
affects populations of rare glade plant species. Experimental and control plots
were established within the glades. Experimental plots were burned in April
2004, and all plots were surveyed during two subsequent growing seasons (2004
and 2005). Populations of three of 14 species of conservation concern declined
significantly after the initial fire but recovered the next year. Among other
herbaceous species, only five and two differed in population size in 2004 and
2005, respectively. In 2004, more species were more abundant in control than
burned plots, but this difference was not detected in 2005. Multivariate
community-level analyses of species presence-absence suggested that the effects
of fire were negligible by the 2005 survey. Populations of young trees that had
invaded the glades declined dramatically as a result of treatment fires. These
results suggest that the reintroduction of fire will not harm glade species and
may help prevent encroachment of the surrounding forest.
longleaf pine forests
affects populations of rare glade plant species. Experimental and control plots
were established within the glades. Experimental plots were burned in April
2004, and all plots were surveyed during two subsequent growing seasons (2004
and 2005). Populations of three of 14 species of conservation concern declined
significantly after the initial fire but recovered the next year. Among other
herbaceous species, only five and two differed in population size in 2004 and
2005, respectively. In 2004, more species were more abundant in control than
burned plots, but this difference was not detected in 2005. Multivariate
community-level analyses of species presence-absence suggested that the effects
of fire were negligible by the 2005 survey. Populations of young trees that had
invaded the glades declined dramatically as a result of treatment fires. These
results suggest that the reintroduction of fire will not harm glade species and
may help prevent encroachment of the surrounding forest.
Which is good, since The
Nature Conservancy of Alabama and its partners have conducted a number
of controlled burns to date at the Kathy
Stiles Freeland Bibb County Glades Preserve (photo at right; note blackened
lower trunks of pines). After seeing the results, I have no doubts that these
prescribed fires have enhanced the vigor and diversity of the herbaceous
component in glade-forest ecotones, and served to reverse the slow
encroachment upon these glades of adjacent woodland communities.
 Considerable recent information about Spigelia gentianoides
can be found in "Spigelia gentianoides, Gentian pinkroot, 5-Year
Review: Summary and Evaluation" (2009), [available at http://www.fws.gov/southeast/5yearReviews/5yearreviews/20090121GentianPinkroot.pdf]
by the U.S. Fish and Wildlife Service,
Panama City,
Considerable recent information about Spigelia gentianoides
can be found in "Spigelia gentianoides, Gentian pinkroot, 5-Year
Review: Summary and Evaluation" (2009), [available at http://www.fws.gov/southeast/5yearReviews/5yearreviews/20090121GentianPinkroot.pdf]
by the U.S. Fish and Wildlife Service,
Panama City, Florida Field Office (Vivian Negrón-Ortiz, Lead Recovery Botanist).
Among the findings reported: there are five locations where the endangered
variety gentianoides is known to be extant, including two sizeable occurrences
on state-owned land in Jackson County, Florida. At Apalachee Wildlife Management
Area (where discovered in 2007), more than1,700 individuals are known, divided
into two subpopulations, and Three Rivers State Park is estimated to support
more than1000 individuals. A third Jackson County population, on private land,
was thought to consist of only four plants. The fourth Florida population, of
about 100 plants, is known from Calhoun County, on a preserve owned by The
Nature Conservancy. The Castanea paper (p. 190) reported a personal
communication from John MacDonald that he had discovered this variety in
adjacent Alabama. This occurrence, in the Geneva State Forest (Geneva County)
was estimated in 2007 to contain about 400 individuals.
Florida Field Office (Vivian Negrón-Ortiz, Lead Recovery Botanist).
Among the findings reported: there are five locations where the endangered
variety gentianoides is known to be extant, including two sizeable occurrences
on state-owned land in Jackson County, Florida. At Apalachee Wildlife Management
Area (where discovered in 2007), more than1,700 individuals are known, divided
into two subpopulations, and Three Rivers State Park is estimated to support
more than1000 individuals. A third Jackson County population, on private land,
was thought to consist of only four plants. The fourth Florida population, of
about 100 plants, is known from Calhoun County, on a preserve owned by The
Nature Conservancy. The Castanea paper (p. 190) reported a personal
communication from John MacDonald that he had discovered this variety in
adjacent Alabama. This occurrence, in the Geneva State Forest (Geneva County)
was estimated in 2007 to contain about 400 individuals.
Regarding variety alabamensis K. Gould, Negrón-Ortiz et
al., using belt transects, estimated there were a total of 3,653 individuals on
4 of the 17 Ketona dolomite glades where it had previously been found.
The recommendations of the USFWS report include the suggestion
that "a taxonomic study using a multi-data approach (e.g., morphology,
molecular studies) is encouraged for discerning whether the two varieties
represent distinct species. If the results suggest that the varieties represent
distinct species, then both should be considered for federal protection." However, the only information available since the
Castanea paper and relevant to this issue that was mentioned in the USFWS report
was the following:
protection." However, the only information available since the
Castanea paper and relevant to this issue that was mentioned in the USFWS report
was the following:
Electrophoretic
studies indicated that the genetic identity between the S. gentianoides
varieties is high. The Florida sample used in the study, presently growing at
the Historic Bok Sanctuary (Sanctuary), from seeds collected at Calhoun County,
included a relatively narrow subset of the genetic variation found in the alabamensis
populations (Affolter 2005*). The majority of allozyme diversity in var. alabamensis
was found to be distributed within rather than among glades, therefore the
author suggested that nearly all the genetic diversity will be captured by
preserving the largest glades.
*
Affolter, J. M. 2005. Conservation biology of Spigelia gentianoides and S.
marilandica: genetic variation, reproduction biology, and propagation. Final
project to the Georgia Cooperative Fish and Wildlife Research Unit. 33
pp.
NOTEWORTHY
ELEMENTS OF THE FLORA:
OTHER RARITIES
At the top of page 195 of the Castanea article I wrote, "It
seems certain that still other botanical (and zoological?) rarities occur on
these glades and await future detection." Some of the insects, at least, of the Ketona Glades have received attention from
zoologists. In 2006 JoVonn G. Hill, of the Mississippi Entomological
Museum at Mississippi State University (Starkville) reported "The First
Record of the Green Stinkbug Chlorochroa persimilis Horvath (Hemiptera:
Pentatomidae) from Alabama" [Marginalia Insecta 1(2):1-2]. The next
year Hill reported on "The Grasshoppers (Orthoptera: Acrididae) of the Bibb
County Glades, Alabama" [Marginalia Insecta 2(1):1-3. 2007].
According to Hill, "The Mississippi Entomological Museum (MEM) visited these glades three
times (1-2 May, 4-6 June, and 6-7 August 2003) for the annual William H. Cross
Collecting Expedition," when nine species of grasshoppers were found. Hill
returned to the glades twice in 2005, on August 13th and on September 8th and 9th.
"These trips yielded 10 additional species of grasshoppers resulting in a
total of 19 species for the site." Hill's 2007 paper continues:
the insects, at least, of the Ketona Glades have received attention from
zoologists. In 2006 JoVonn G. Hill, of the Mississippi Entomological
Museum at Mississippi State University (Starkville) reported "The First
Record of the Green Stinkbug Chlorochroa persimilis Horvath (Hemiptera:
Pentatomidae) from Alabama" [Marginalia Insecta 1(2):1-2]. The next
year Hill reported on "The Grasshoppers (Orthoptera: Acrididae) of the Bibb
County Glades, Alabama" [Marginalia Insecta 2(1):1-3. 2007].
According to Hill, "The Mississippi Entomological Museum (MEM) visited these glades three
times (1-2 May, 4-6 June, and 6-7 August 2003) for the annual William H. Cross
Collecting Expedition," when nine species of grasshoppers were found. Hill
returned to the glades twice in 2005, on August 13th and on September 8th and 9th.
"These trips yielded 10 additional species of grasshoppers resulting in a
total of 19 species for the site." Hill's 2007 paper continues:
No new state
records or new species were found in these habitats, but there were a few
interesting finds. Both Spharagemon cristatum (Scudder) and S.
marmorata picta (Scudder) are species that are typically associated with
sandy habitats (Dakin and Hayes, 1970 and Otte 1984). Here, they were found
dwelling on the rocky dolomite outcrops on the most extensive glade on a bluff
overlooking the Little Cahaba River. (Figure 1*). Although S. cristatum
is known to occupy sandy areas near rivers (there were some sandy margins on the
river below), there were significant numbers of this species occupying the rocky
outcrops. Both Spharagemon species were camouflaged quite well when
resting on the exposed dolomite. Another interesting find was that of Melanoplus
impudicus Scudder, which can be found in dry areas of sparse vegetation on
rocky or sandy areas (Gurney, 1941 and Dakin and Hayes, 1970). Gurney (1941)
figured a shale barren habitat at Short Mt. near Mt. Jackson, Massanutten Mts.
Virginia where M. impudicus was found and this shale barren is similar in
appearance to the Bibb County Glades.
* Reproduced at
right, with a Pinus palustris that I recognize as the leftmost in a grouping
of that species in a photo featured elsewhere on this web site (http://www.jimbotany.com/pinuspal.jpg), and marking the
site to me as the one I named "South Goat Glade."
Hill's 2006 paper is available at http://mississippientomologicalmuseum.org.msstate.edu//MEM.Pubs/Marginalia.Insecta/2006.papers/Marginalia.Insecta.Vol.1.2.pdf,
the 2007 paper at http://mississippientomologicalmuseum.org.msstate.edu/MEM.Pubs/Marginalia.Insecta/2007.papers/Marginalia.insecta.Vol.2.1.pdf.
ERRATA
Page 157, first paragraph: replace "Psora rubiformis
(Ach.) Hooker vel aff." with "Psora pseudorussellii Timdal"
(I have re-identified my collection, which is apparently yet another range
extension (according to the Consortium of North American Lichen Herbaria,
previously recorded from no closer than Arkansas; see http://symbiota.org/nalichens/index.php).
Pages 163, 168, 178 (twice), 182, 186, 196: correct the spelling
of "Brown's" Dam to "Browne's" Dam, as this
historic site was named for William P. Browne, according to Bibb County,
Alabama: The First Hundred Years, by Rhoda C. Ellison (The University of
Alabama Press, 1984).
Page 169:
first line of third paragraph: delete "a decumbent habit,"
Page
191, end of third paragraph: delete ", UNA"
Page
195,
line 11:
delete "(Muhl. ex Willd.)"
Page
199, under ASTERACEAE, in text for Coreopsis grandiflora var. inclinata
X C. pubescens,
replace "A. & S. 6719"
with "A. 11927"
Page
200, under CONVOLVULACEAE: the correct spelling of the specific epithet
of the Calystegia
is
apparently "catesbeiana"
Page
200, under CYPERACEAE, in text for Rhynchospora globularis var. pinetorum:
replace "(Britt. & Small ex Small) Gale" with "(Britt. &
Small) Gale"
Page 201, under
FAGACEAE, correct "Quercus margarettiae" to "Quercus
margarettae" [an error copied from USDA 2000 (PLANTS
Database, since corrected there)].
Page
201, line 11: add "(AUA)" after "7724"
Page
202,
line 43:
replace "A. and S. 7413"
with "A. and S. 7430"
Page
202, line 45: replace "J.
Allison and C. Oberholster 7049"
with
"A. and S. 7413b"
ADDENDA
These are mostly additions to the flora or are taxonomic
changes (made subsequent to going to press, or then somewhat controversial but
since gaining wider acceptance by systematists). I cannot claim to have kept up
with all of these; most of the credit must go to Alan Weakley, via his "Flora").
Page
154:
the first author's current address for correspondence: 2242 Highway 212 SW;
Conyers,
Georgia 30094.
Page 159, end of second paragraph: the URL for the referenced
web site has changed to http://www.jimbotany.com/.
Page 199, before AGAVACEAE add "ADOXACEAE" and list under it "Viburnum
rufidulum Raf." (transfer this from CAPRIFOLIACEAE. page 200).
Page 199, under APIACEAE, Thaspium barbinode var. chapmanii:
add "[T. chapmanii (Coult. & Rose) Small]" as this taxon
may reasonably be treated as a full species (e.g., 2010 edition of
"Weakley's Flora").
Page 199: transfer
taxa listed under ASCLEPIADACEAE to APOCYNACEAE.
Page 199, under APOCYNACEAE (ASCLEPIADACEAE): the correct name
for the "Matelea" is now regarded to be Gonolobus suberosus
(L.) R. Brown.
Page 199, under ASTERACEAE, Acmella oppositifolia var. repens:
add "[A. repens (Walt.) L.C.
Rich. in Pers.], as this taxon may reasonably be treated as a full
species (e.g., 2010 edition of "Weakley's Flora").
Page 199, under ASTERACEAE, after "Coreopsis major
Walt." add the clarifying "var. major"
Page 199, under ASTERACEAE, Silphium astericus var. angustatum,
add "[S. asteriscus var. dentatum (Ell.) Chapm.]" as the
former taxon may be considered synonymous with the latter (e.g., Flora
of North America North of Mexico, Vol. 19).
Page 199, under ASTERACEAE, Silphium trifoliatum var. latifolium,
add "[Silphium asteriscus L. var. latifolium (A. Gray)
J.A. Clevinger]" as this taxon is so treated in Flora
of North America North of Mexico, Vol. 19.
Page
200, under BRASSICACEAE, Arabis canadensis: add "[Boechera
canadensis (L.) Al-Shehbaz], as there is now strong evidence for placing most southeastern
"Arabis" spp. in Boechera
Löve
& Löve.
Page 200, after
BUDDLEJACEAE, add "[TETRACHONDRACEAE]", as Buddleja seems to
belong to the Scrophulariaceae (so no Buddlejaceae), and while the
placement of Polypremum procumbens remains controversial, it is perhaps
best placed with Tetrachondra Petrie [Oxelman,
B., M. Backlund, and B. Bremer. 1999. Relationships of the Buddlejaceae s.l.
investigated using parsimony jackknife and branch support
analysis of chloroplast ndhF and rbcL sequence data. Syst. Bot. 24: 164-182].
Page
200, under CAMPANULACEAE: add "Triodanis
perfoliata (L.) Nieuw."
Page 200, under
CARYOPHYLLACEAE: at end of column 1 add "[S.
caroliniana Walt. var. wherryi (Small)
Fern.]".
Page 200,
CLUSIACEAE: see first note below for page 201.
Page 200, under
CONVOLVULACEAE: delete "(identification tentative)" from the Calystegia.
Page
200, under CYPERACEAE, Rhynchospora globularis var. pinetorum:
after "Gale" insert "[R. pinetorum Britton & Small]".
Page
200, under CYPERACEAE: Scleria
verticillata, delete "(single
locality)".
Page
200, under EUPHORBIACEAE, transfer the Leptopus to PHYLLANTHACEAE after
adding this family name before PLANTAGINACEAE (page 202).
Page 201, before
IRIDACEAE, add "HYPERICACEAE" and transfer the Hypericum spp.
from page 200 under CLUSIACEAE (delete the latter family). After Hypericum
densiflorum text add "[referable to the narrow-leaved plants named as H.
interior Small, probably a good subspecies but the combination apparently
not yet made]".
Page 201, before
IRIDACEAE, add "HYPOXIDACEAE" and transfer the Hypoxis from the
former family to the latter.
Page 201, under
LILIACEAE: Aletris is now regarded as belonging in NARTHECIACEAE (insert
this family before NYCTAGINACEAE, page 202, Allium and Nothoscordum
in the ALLIACEAE (insert before ANACARDIACEAE, page 198), Camassia and Schoenolirion
in the AGAVACEAE (page 198). Polygonatum has been placed in the RUSCACEAE
(insert before RUTACEAE, page 203), Trillium in the TRILLIACEAE (insert
before ULMACEAE, page 203), and Uvularia in the COLCHICACEAE
(insert before COMMELINACEAE, page 200). The three
species listed as Melanthium have been transferred to Veratrum, in
the MELANTHIACEAE (insert this family after MALVACEAE, page 201. In the entry
for M. latifolium, after "Walt." insert ", Veratrum
latifolium (Desr.) Zomlefer" and in the entry for M. virginicum,
after "L.}" insert "[Veratrum virginicum (L.)
Ait.]".
Page 201, under LOGANIACEAE: Gelsemium has been made the
type-genus of its own family; insert "GELSEMIACEAE" before
GENTIANACEAE and transfer G. sempervirens.
Page 202, under POACEAE: after Sporobolus vaginiflorus
insert "Tridens chapmanii (Small)
Chase" and place the combination "Tridens flavus var. chapmanii"
(with the corresponding authorities) in brackets as a synonym.
Page 202, under PRIMULACEAE: Dodecatheon L. is now
regarded as insufficiently distinct from Primula L.; insert the new
combination "Primula meadia (L.) A.R. Mast & Reveal" and
place Dodecatheon meadia L. in brackets as a synonym.
Page 202, under PRIMULACEAE: Lysimachia is now regarded
as belonging to the MYRSINACEAE; insert this name following MYRICACEAE (page
201) and transfer the Lysimachia spp. there.
Page
202, under RANUNCULACEAE: add "Thalictrum
revolutum DC."
Page
203,
under
ROSACEAE: add "Prunus
angustifolia Marsh.
(single
locality)".
Page
203,
under
RUBIACEAE: add "Houstonia
canadensis Willd."
Page 203, under
SAXIFRAGACEAE: the only genus listed, Parnassia, is now (again) regarded
as the type genus of its own family; insert "PARNASSIACEAE" before
PASSIFLORACEAE, transfer P. grandifolia to the latter, and delete "SAXIFRAGACEAE."
Page 203, under
SCROPHULARIACEAE: the genera Agalinis, Aureolaria, Buchnera,
Castilleja, Pedicularis, and Seymeria
are now regarded as members of the Orobanchaceae (page 202); make the necessary
transfers. Leucospora, Mecardonia, Penstemon, Veronica, and Veronicastrum
are now placed in the Plantaginaceae (page 202); make these transfers as well,
and delete "SCROPHULARIACEAE."
Page 203, under
ULMACEAE: Celtis L. is probably not a member of the Ulmaceae, but I defer
placing it in another family (e.g. Celtidaceae, Cannabaceae) pending
clarification from additional studies.
Page 203, under
VERBENACEAE: Callicarpa L. may better be regarded as a member of the
Lamiaceae, but I defer making this transfer pending further studies (which may
support a "Callicarpaceae").
Page 203, under XYRIDACEAE: after "Xyris tennesseensis
Kral" insert "[X. spathifolia Kral & Moffett]"
(described in J. Bot. Res. Inst. Texas 3:469–478. 2009 and available at this
writing for download at http://www.brit.org/fileadmin/Publications/JBotResInstTexas_3_2/469-478_KralMoffett_Xyris_JBRIT3_2__01.pdf). Unlike the other
Ketona Glade endemics, this entity is known to date from a solitary occurrence,
where it grows in association with the closely related X. tennesseensis.
Until an additional natural occurrence is discovered, it seems best regarded as
a mutant derived in situ from its intimate associate.
Although multiple distinguishing features are cited, these are of a kind to
suggest pleiotropy (a single developmental gene affecting several distinct and
seemingly unrelated phenotypic characteristics). Sequencing
the two xyrids would be helpful in testing this hypothesis. In the
meantime, I hope that the publication results in site protection and
conservation management, as well as other ex situ cultivation efforts in
addition to
those mentioned in the paper, so that the mystery of the evolution of "Xyris
spathifolia" can be unraveled before this
interesting phenotype becomes extinct.
Page 205,
bibliographic entry beginning "USDA": the PLANTS Database has since
changed its URL to http://plants.usda.gov/.
As more is
learned about the
flora
of the Ketona Dolomite Glades (or additional errata detected in the Castanea
paper) this page will be updated.
| Home |
Castanea
paper Table of Contents |
 |
 |
|
Most recent update:
September 30, 2012
 Liatris oligocephala
is the only known Liatris lacking
tiny bristles on the ribs of its achenes. It is also the only Liatris
that
produces mostly one to
Liatris oligocephala
is the only known Liatris lacking
tiny bristles on the ribs of its achenes. It is also the only Liatris
that
produces mostly one to
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}