![]()
 Another
conspicuous and unfamiliar species observed on glades during the initial canoe
expedition was a leafy-stemmed, opposite-leaved herbaceous plant,
clearly still some weeks from
flowering, whose stem and leaves were so densely beset
Another
conspicuous and unfamiliar species observed on glades during the initial canoe
expedition was a leafy-stemmed, opposite-leaved herbaceous plant,
clearly still some weeks from
flowering, whose stem and leaves were so densely beset with
gland-tipped hairs that they gave it a
sticky feel. Its overall look was suggestive of a Silphium,
but consultation of readily available
literature (e.g., Perry 1937, Cronquist 1980) gave no
indication that any taxon so copiously glandular
occurred in that genus. In a few weeks the plants began to flower and
both their generic placement
(Silphium) and their distinctiveness were manifest.
with
gland-tipped hairs that they gave it a
sticky feel. Its overall look was suggestive of a Silphium,
but consultation of readily available
literature (e.g., Perry 1937, Cronquist 1980) gave no
indication that any taxon so copiously glandular
occurred in that genus. In a few weeks the plants began to flower and
both their generic placement
(Silphium) and their distinctiveness were manifest.
Silphium glutinosum J. Allison, sp. nov. TYPE: Alabama: Bibb County, ca. 17.8 km NE of Centreville, ca. 2 km NNW of the mouth of Alligator Creek. "Alligator Glade," Ketona Dolomite outcrop ca. 0.2 km W of Alligator Creek, 19 Aug 2000, James R. Allison 12503 (holotype, GH; isotypes: AUA, DUKE, FLAS, FSU, GA, IBE, JSU, MICH, MO, NCU, NY, TAMU, UARK, UNA, US, USCH, VDB). Figures 11 and 12.
Species a congeneris combinatione
capitulorum radiis vulgo tredecim cum indumento denso
pro  parte maxima glandulifero caulium et foliorum et involucrorum haud
aegre distinguitur.
parte maxima glandulifero caulium et foliorum et involucrorum haud
aegre distinguitur.
Perennial herb,
fleshy-rooted from a short rhizome or nearly erect caudex, vegetative
portion
densely pubescent
throughout with a mixture of long, stipitate-glandular hairs and mostly
shorter,
pointed, eglandular hairs, somewhat resinous-aromatic.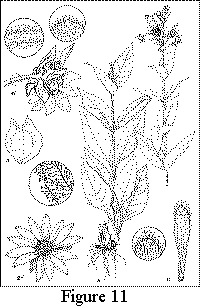 Stems
1-several, (3.6) 8-15 (18) dm tall,
unbranched except for
inflorescence, (4) 5.5-10 (11) mm thick at base, finely sulcate,
terete,
yellowish green or sometimes maroon-tinted, especially near the base,
nodes with persistent leaves
at anthesis (4) 6 or 7 (10) below the branching of the inflorescence. Leaves
opposite or rarely
alternate, very rarely 3 at a node, dull, upper surface dark yellowish
green, lower surface paler,
margins ciliate; the lowest (at anthesis) the largest, ovate or
lanceolate, larger blades 15-22 (24) cm
long, 7-12 cm wide, coarsely and shallowly toothed, particularly toward
the base, with winged
petioles 6-15 (16) cm long, pubescent and ciliate like the blade (often proximally with longer,
eglandular hairs); leaves upwardly gradually reduced in size, with
shorter and more broadly winged
petioles, becoming sessile and entire and grading into ovate or
lanceolate (-broadly elliptic) bracts
in the inflorescence. Inflorescence of (1) several to many
(>30) heads in an open, broad (to 36 cm)
panicle, the branches stiffish, the central peduncle of a main branch
6-13 (15) cm long (laterals
shorter), straight (or curved early in anthesis and the heads nodding),
bracteolate at the base or
sometimes above the middle; involucre
Stems
1-several, (3.6) 8-15 (18) dm tall,
unbranched except for
inflorescence, (4) 5.5-10 (11) mm thick at base, finely sulcate,
terete,
yellowish green or sometimes maroon-tinted, especially near the base,
nodes with persistent leaves
at anthesis (4) 6 or 7 (10) below the branching of the inflorescence. Leaves
opposite or rarely
alternate, very rarely 3 at a node, dull, upper surface dark yellowish
green, lower surface paler,
margins ciliate; the lowest (at anthesis) the largest, ovate or
lanceolate, larger blades 15-22 (24) cm
long, 7-12 cm wide, coarsely and shallowly toothed, particularly toward
the base, with winged
petioles 6-15 (16) cm long, pubescent and ciliate like the blade (often proximally with longer,
eglandular hairs); leaves upwardly gradually reduced in size, with
shorter and more broadly winged
petioles, becoming sessile and entire and grading into ovate or
lanceolate (-broadly elliptic) bracts
in the inflorescence. Inflorescence of (1) several to many
(>30) heads in an open, broad (to 36 cm)
panicle, the branches stiffish, the central peduncle of a main branch
6-13 (15) cm long (laterals
shorter), straight (or curved early in anthesis and the heads nodding),
bracteolate at the base or
sometimes above the middle; involucre  shallowly
campanulate, 3-4.3 cm(7)
broad, the phyllaries in
about 4 series, mostly 19-23, 11-19 mm long and 5.5-11 mm wide,
membranaceous, rather loosely
overlapping, ribbed proximally, the outermost usually the longest,
ascending or sometimes (especially
those of the larger, central head of an inflorescence-branch),
spreading and with margins and tips
somewhat recurving, pubescent and ciliate like the leaves, lanceolate
and acute, inwardly
progressively reduced and passing through ovate to elliptic and obtuse
and then oblanceolate with
a broadly rounded apex, the innermost ca. 8 mm X 2 mm,
the pubescence progressively more
confined to the distal, dilated portion and the ribbed proximal area
becoming proportionately greater; pales 9-11 mm long, linear-oblanceolate, apices distinctly
obtuse, with both glandular and eglandular,
scalelike pubescence; rays (8) 12-14 (16), usually 13, (1.7) 2.0-2.7 cm
long, linear-elliptic, shallowly
emarginate, pale yellow; disk mostly 1.4-2.2 cm broad, the numerous
(>60) corollas tubular, greenish
yellow, 6-7 mm long, with ascending, broadly triangular lobes. Fruit
a broadly ovate to orbicular
achene, 6-8.5 mm long (excluding pappus teeth), with wing ca. 1.5
shallowly
campanulate, 3-4.3 cm(7)
broad, the phyllaries in
about 4 series, mostly 19-23, 11-19 mm long and 5.5-11 mm wide,
membranaceous, rather loosely
overlapping, ribbed proximally, the outermost usually the longest,
ascending or sometimes (especially
those of the larger, central head of an inflorescence-branch),
spreading and with margins and tips
somewhat recurving, pubescent and ciliate like the leaves, lanceolate
and acute, inwardly
progressively reduced and passing through ovate to elliptic and obtuse
and then oblanceolate with
a broadly rounded apex, the innermost ca. 8 mm X 2 mm,
the pubescence progressively more
confined to the distal, dilated portion and the ribbed proximal area
becoming proportionately greater; pales 9-11 mm long, linear-oblanceolate, apices distinctly
obtuse, with both glandular and eglandular,
scalelike pubescence; rays (8) 12-14 (16), usually 13, (1.7) 2.0-2.7 cm
long, linear-elliptic, shallowly
emarginate, pale yellow; disk mostly 1.4-2.2 cm broad, the numerous
(>60) corollas tubular, greenish
yellow, 6-7 mm long, with ascending, broadly triangular lobes. Fruit
a broadly ovate to orbicular
achene, 6-8.5 mm long (excluding pappus teeth), with wing ca. 1.5 mm
wide at the base of the sinus,
the body appressed white-hairy and ciliate in the sinus,
otherwise smooth or sparsely pale-strigillose,
gray-brown when ripe. Pappus of two teeth 0.5-2.0 (3) mm
long, rarely one or two smaller, narrower
secondary awns persisting. Chromosome number 2n = 14 (A. and S. 8107,
UNA).
mm
wide at the base of the sinus,
the body appressed white-hairy and ciliate in the sinus,
otherwise smooth or sparsely pale-strigillose,
gray-brown when ripe. Pappus of two teeth 0.5-2.0 (3) mm
long, rarely one or two smaller, narrower
secondary awns persisting. Chromosome number 2n = 14 (A. and S. 8107,
UNA).
 Flowering June-October, fruiting
July-frost.
Flowering June-October, fruiting
July-frost.
English Name: Sticky Rosinweed.
Paratypes. Alabama: Bibb
Co., 10.0 km NNE of Centreville, rocky bank of Little Schultz Cr.,
15 Aug 1998, J. Allison and M. Moffett 11326 (AUA); 12.1 km
NNE of Centreville, "Lady-tresses
Glade South," 22 Aug 1999, A. 12170 (AUA, GH, NY, UNA, US);
13.5 km NE of Centreville, Six
Mile Cr., "The Sinks," 15 Aug 1998, J. Allison and M. Moffett 11324
(GA); 14.4 km NE of
Centreville, "Beaver Glade," 11 Oct 1992, A. and S. 7308
(UNA); 14.6 km NE of Centreville, "Goat
Glade South," 8 Oct 2000, A. 12564 (AUA, GA, GH, JSU,
UNA, US, VDB); 14.9 km NE of
Centreville, "Brown's Dam North Glade West," 16 Aug 1998, A. 11329
(GA, GH, IBE, JSU, MICH,
MO, NCU, NY, TAMU, UNA, US, VDB); 15.3 km NE of Centreville, "Desmond's
Glade," 22 Jul
2000, A. 12445 (AUA, DUKE, FLAS, FSU, GA, GH, IBE, JSU,
MICH, MO, NY, TAMU, TENN,
UARK, UNA, US, USCH, VDB).
South," 8 Oct 2000, A. 12564 (AUA, GA, GH, JSU,
UNA, US, VDB); 14.9 km NE of
Centreville, "Brown's Dam North Glade West," 16 Aug 1998, A. 11329
(GA, GH, IBE, JSU, MICH,
MO, NCU, NY, TAMU, UNA, US, VDB); 15.3 km NE of Centreville, "Desmond's
Glade," 22 Jul
2000, A. 12445 (AUA, DUKE, FLAS, FSU, GA, GH, IBE, JSU,
MICH, MO, NY, TAMU, TENN,
UARK, UNA, US, USCH, VDB).
 In contrast with the other Ketona
Glade endemics, Silphium glutinosum seems most vigorous
in partial shade, though it will grow in full sun of the open glade.
Perhaps it is no coincidence
that it
is also the only endemic taxon of the Ketona Dolomite glades that does
not appear to be completely
restricted to them. Besides on glades it can sometimes be found in
rocky places along streams and
occasionally along logging roads in the glade region. In view of this,
it is somewhat surprising that
it is absent from several glades occurring near the eastern and western
extremities of the Ketona
Formation in Bibb County.
In contrast with the other Ketona
Glade endemics, Silphium glutinosum seems most vigorous
in partial shade, though it will grow in full sun of the open glade.
Perhaps it is no coincidence
that it
is also the only endemic taxon of the Ketona Dolomite glades that does
not appear to be completely
restricted to them. Besides on glades it can sometimes be found in
rocky places along streams and
occasionally along logging roads in the glade region. In view of this,
it is somewhat surprising that
it is absent from several glades occurring near the eastern and western
extremities of the Ketona
Formation in Bibb County.
Both the gradually reduced leaves, and heads usually with 13 rays(8)
align this taxon with the
Silphium asteriscus complex (e.g., S. asteriscus
L., S. trifoliatum L. and their varieties). Like most
Silphium species, S. glutinosum will sometimes
hybridize with related species(9)
when they grow
nearby. The most common Silphium of roadsides and![Silphium trifoliatum var. latifolium, Bibb County, Alabama [=S. asteriscus var. latifolium]. Note the well-spaced nodes as compared with S. glutinosum.](SIASLATI.JPG) thin woods
in the vicinity of the Ketona Glades
is S. trifoliatum L. var. latifolium Gray.
Where the glade-forest ecotone had been disturbed by
logging or road construction, plants of apparently intermediate
morphology were sometimes found
(e.g., A. and S. 7301, UNA; A. and S. 7869, VDB).
Such plants were also found in rocky areas along
major streams of the Ridge and Valley in Bibb County, and were often
more abundant than either
parental species (e.g., A. and S. 8022, JSU).
thin woods
in the vicinity of the Ketona Glades
is S. trifoliatum L. var. latifolium Gray.
Where the glade-forest ecotone had been disturbed by
logging or road construction, plants of apparently intermediate
morphology were sometimes found
(e.g., A. and S. 7301, UNA; A. and S. 7869, VDB).
Such plants were also found in rocky areas along
major streams of the Ridge and Valley in Bibb County, and were often
more abundant than either
parental species (e.g., A. and S. 8022, JSU).
Commonly, Silphium trifoliatum var. latifolium is essentially glabrous, but Bibb County
populations, otherwise seemingly typical of the variety, usually have
some spreading, transparent,
hairs on the peduncles. The presence of such hairs on the putative ![Silphium glutinosum X S. trifoliatum [asteriscus] var. latifolium](SIGXATRI.JPG) hybrids, intermixed with shorter,
glandular hairs, in combination with intermediacy of leaf shape,
petiole length, and involucre
morphology, seems proof that such plants are indeed of hybrid origin.
hybrids, intermixed with shorter,
glandular hairs, in combination with intermediacy of leaf shape,
petiole length, and involucre
morphology, seems proof that such plants are indeed of hybrid origin.
Another, much less common putative hybrid, we observed where the parental taxa grew in close proximity, was Silphium asteriscus L. var. asteriscus X S. glutinosum (e.g., A. and S. 8474, UNA; A. and S. 8475, VDB). As many as three genomes seem to have been involved in the formation of one plant seen along Little Schultz Creek, apparently S. asteriscus L. var. angustatum Gray X S. trifoliatum var. latifolium X S. glutinosum. Other putative hybrids of Silphium but not involving S. glutinosum were also seen, such as S. compositum Michx. X S. laciniatum L. (A. 12508, UNA). As all published chromosome counts in Silphium are diploid (Settle and Fisher 1970, Cronquist 1980, subsequent volumes of Index to Plant Chromosome Numbers), and given the frequency of hybridization in human-perturbed habitats, it appears that, among related taxa of Silphium, barriers to gene flow are chiefly ecological. Alteration of the landscape by humans appears to have disrupted these barriers, just as in Coreopsis.
Of course, taxa with overlapping
periods of anthesis but different habitat preferences came into
contact occasionally prior to widespread habitat manipulation by
humans. It appears that under
certain conditions, resulting hybrids may have outcompeted the parental
species and founded
populations that, over time, became stabilized in their morphology and
appear now as fully
independent taxa. One possible such taxon, of Tennessee and Kentucky,
has been called Silphium
integrifolium Michx. var. gattingeri Perry. Kral (1983)
opined that these plants suggested a blending
of S. integrifolium and S. trifoliatum var. latifolium.
Upon examining specimens at VDB
the rightness of Kral's interpretation seemed compelling, the large
heads with many rays suggesting S.
integrifolium, the petiolate lower leaves and smooth stems
suggesting in turn S. trifoliatum var.  latifolium. The lack of intermediacy in pubescence could be
explained by the loss of integrifolium
alleles in recombination, or recessiveness of integrifolium pubescence
alleles. A hybrid origin for
taxon gattingeri has not yet been proven, but support for
this hypothesis is provided by a seemingly
analogous situation in Alabama.
latifolium. The lack of intermediacy in pubescence could be
explained by the loss of integrifolium
alleles in recombination, or recessiveness of integrifolium pubescence
alleles. A hybrid origin for
taxon gattingeri has not yet been proven, but support for
this hypothesis is provided by a seemingly
analogous situation in Alabama.
After
showing Robert Kral Silphium glutinosum in the field in Bibb
County, he called our
attention to densely glandular specimens labeled S. integrifolium
he had collected from the Black
Belt
province of Alabama, in Dallas County (Kral 48820, VDB) and
Perry County (Kral 47891, VDB).
The authors visited both of these populations, and "windshield surveying" along highways
and back
roads resulted in the discovery of well over a dozen additional
populations of a densely glandular
Silphium in those two counties. After comparison of living
populations and preserved material of
these plants with S. integrifolium and with S.
glutinosum, we determined that a suite of characters
exists that distinguish the Dallas and Perry County plant from either
of those species.
labeled S. integrifolium
he had collected from the Black
Belt
province of Alabama, in Dallas County (Kral 48820, VDB) and
Perry County (Kral 47891, VDB).
The authors visited both of these populations, and "windshield surveying" along highways
and back
roads resulted in the discovery of well over a dozen additional
populations of a densely glandular
Silphium in those two counties. After comparison of living
populations and preserved material of
these plants with S. integrifolium and with S.
glutinosum, we determined that a suite of characters
exists that distinguish the Dallas and Perry County plant from either
of those species.
 The
number of ray flowers consistently greater than 13 would cause these plants to
key to Silphium
integrifolium in, for example, Cronquist (1980), supporting Kral's
determinations of his Dallas and
Perry County collections. Plants truly conforming to that species,
however, are known in Alabama
only from two counties (Pickens and Sumter) on the Mississippi border,
localities at least 80 km to
the northwest of the glandular Dallas and Perry County populations.
The
number of ray flowers consistently greater than 13 would cause these plants to
key to Silphium
integrifolium in, for example, Cronquist (1980), supporting Kral's
determinations of his Dallas and
Perry County collections. Plants truly conforming to that species,
however, are known in Alabama
only from two counties (Pickens and Sumter) on the Mississippi border,
localities at least 80 km to
the northwest of the glandular Dallas and Perry County populations.
Silphium integrifolium differs from S. glutinosum in many respects. The former is, under favorable conditions in the wild and in the garden (e.g., garden of Allison), a taller plant with nodes slightly closer together and therefore more numerous. Those leaves that persist until anthesis in S. integrifolium are normally sessile and, like the stem, eglandular. It also has a narrower inflorescence, on average with fewer but larger heads, these with consistently more numerous rays and phyllaries and with acute receptacular bracts (pales or chaff). The characters of lower leaf petiole length, plant height, and node number are evident in the field but not in the herbarium, as tall species such as S. integrifolium are virtually always represented there only by the inflorescence and a very few upper nodes ("top snatched"); the specimen shown in Settle and Fisher (1970) is typical in this regard.
|
|
|
|
|
Plants of the Dallas and Perry County Silphium are intermediate between S. glutinosum and S. integrifolium for some characters and for others resemble one or the other of those species (Table 2). It would be less than satisfactory to treat them as S. glutinosum X S. integrifolium, when they are allopatric with respect to both of those species and therefore have a present existence and future evolution independent of both, not to mention that the Dallas-Perry entity is only hypothetically of hybrid origin. To choose the alternative of making it a new variety (or subspecies) would require selecting one or the other putative parental species as being of closer affinity, when there is, at least at present, no clear basis for choosing between them. As these populations are consistent in their morphology, are readily distinguished from all previously described taxa, and are sympatric with neither of the species they most closely resemble, they are described here as a new species, Silphium perplexum.
|
Table 2: Morphological comparison of three Silphium taxa |
S. glutinosum |
S. perplexum |
S. integrifolium |
|
plant height |
mostly 8-15 dm |
mostly 15-20 dm |
often >15 dm |
|
nodes below inflorescence |
mostly 6 or 7 |
mostly 9-12 |
often 10 or more |
|
stem and leaf glandular hairs |
long |
short |
absent |
|
lowest petiole length at anthesis |
mostly 6-15 cm |
mostly 5-8 cm |
mostly 0 cm |
|
inflorescence |
open |
open or ± congested |
± congested |
|
chaff apices |
obtuse |
obtuse to acutish |
strongly acute |
|
number of rays per head |
(8) 12-14 (16) |
(17) 19-23 (33) |
(16) 21-34 (35) |
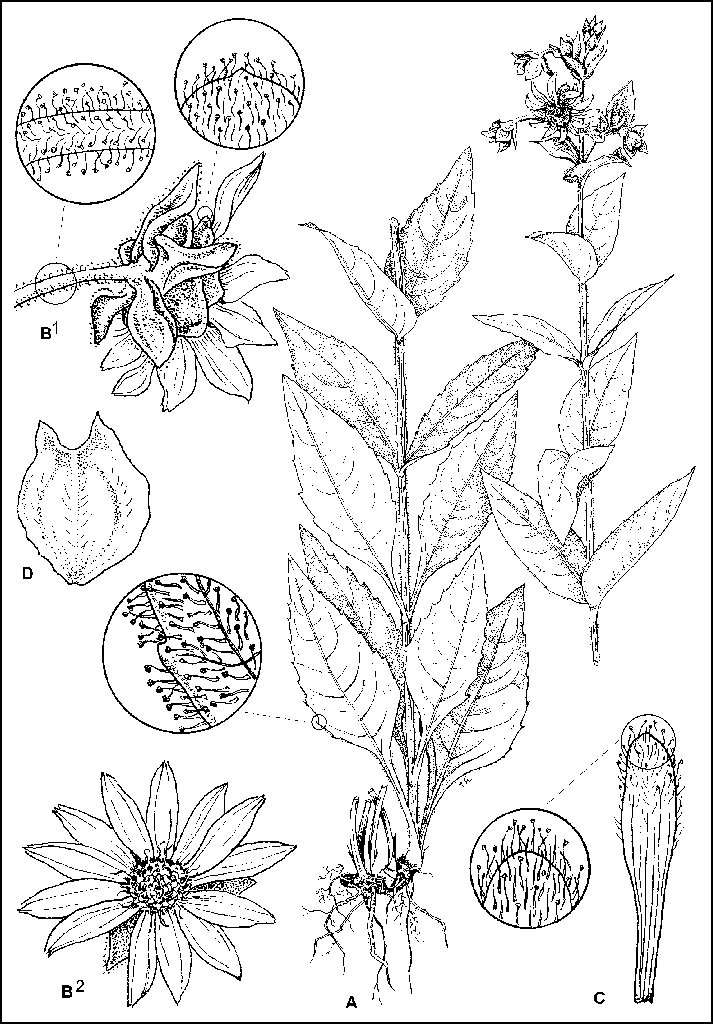
Silphium perplexum J. Allison, sp. nov. TYPE: Alabama: Dallas
County, ca. 15.5 km SW of Selma
and ca. 2.4 km WSW of Old Cahaba; S side Co. Rd. 2 at jct. Co. Rd. 9; prairielike openings over
chalk, abundant, 18 Aug 1999, James R. Allison 12153
(holotype, GH; isotypes: AUA, DUKE, FLAS, FSU, GA, JSU, MICH, MO, NY, UNA, US,
VDB). Figure 12.
prairielike openings over
chalk, abundant, 18 Aug 1999, James R. Allison 12153
(holotype, GH; isotypes: AUA, DUKE, FLAS, FSU, GA, JSU, MICH, MO, NY, UNA, US,
VDB). Figure 12.
A S. glutinoso J. Allison caule et foliis et involucris glanduliferis accedit, sed radiis semper plus numerosis (plus quam 16) et caule pilis glandiferis brevioribus, et petiolis plerumque brevioribus recedit.
Similar to Silphium glutinosum in having glandular
hairs on the stem, leaves, involucres, and
pales, but allopatric and growing over chalk rather than dolomite or
limestone, differing
morphologically as follows: stems averaging taller, mostly
15-20 (24.4) dm, to 13 mm thick at base; nodes
usually more numerous, (7) 9 -12 (17); glandular hairs of herbage
averaging distinctly shorter, about
equal in length to the accompanying pointed hairs. Leaves more
often lanceolate and upwardly more
gradually reduced, l arger blades as much as 30 cm long and 15 cm wide,
petioles averaging shorter,
at anthesis the longest at most 5-8 (10) cm long. Heads
usually a little larger, involucre to 4.6 cm
broad, the phyllaries more numerous, mostly 19-32 (35), 8-16 mm long,
slightly more chartaceous,
the outermost slightly shorter than the next interior, rays more
numerous, (17) 19-23 (33), and a
deeper yellow; pales with obtuse or acutish apices. Chromosome number:
2n = 14 (A. and S. 8119,
GH, UNA).
arger blades as much as 30 cm long and 15 cm wide,
petioles averaging shorter,
at anthesis the longest at most 5-8 (10) cm long. Heads
usually a little larger, involucre to 4.6 cm
broad, the phyllaries more numerous, mostly 19-32 (35), 8-16 mm long,
slightly more chartaceous,
the outermost slightly shorter than the next interior, rays more
numerous, (17) 19-23 (33), and a
deeper yellow; pales with obtuse or acutish apices. Chromosome number:
2n = 14 (A. and S. 8119,
GH, UNA).
Flowering July-October, fruiting August-frost.
English Name: Old Cahaba Rosinweed.
Paratypes. Alabama: Dallas Co., just N of Old Cahaba, chalk hills, 6 Oct 1972, R. Kral 48820 (VDB); 19.8 km WNW of Selma, Co. Rd. 45, 14 Jul 1999, J. Allison and A. Schotz 12080 (NY); 20.3 km WNW of Selma, Co. Rd. 88, 18 Aug 1999, A. 12155 (AUA, GH, JSU, NY, UNA, US). Perry Co., dry roadside thickets near Marion, 1 Sep 1885, J. D. Smith s.n. (GH); S of Marion, Ala. Hwy. 5, just N of Washington Cr., prairie remnant, 29 Jul 1972, R. Kral 47891 (VDB); 11.9 km SSE of Marion, Co. Rd. 12, 19 Aug 1999, A. 12161 (AUA, GA, GH, JSU, UNA); 13.4 km SSW of Marion, Co. Rd. 15, 8 Oct 2000, A. 12583 (AUA, GA, GH, JSU, MICH, MO, NY, UNA, US, VDB); 14.5 km SE of Marion, Co. Rd. 6, 19 Aug 1999, A. 12160 (GH, UNA, USCH).
All of the known Silphium perplexum populations are within 13.5 km of the Cahaba River. The type locality is only about 1.3 km from that river, and about 76 km south of and downstream from a riverside population of the Bibb County endemic, S. glutinosum (of course, with meanders the actual length of river is considerably greater). It is easy to envision propagules of the latter being transported downstream by flood waters at a time when the range of S. integrifolium extended somewhat eastward of its present known extent, facilitating hybridization between two formerly allopatric species. The fact that S. perplexum extends over a greater geographic area than one of its putative ancestors, S. glutinosum (ca. 32.4 km across vs. 11.5 km), argues against its formation in the immediate past. In any event, it is certain that its formation preceded 1885 (see paratypes).
One factor that appears to have favored Silphium perplexum over many competing species is unpalatability to cattle. Where prairie habitats within its range have been subjected to grazing—and this would appear to be most, if not all of them—S. perplexum becomes very abundant, sometimes virtually the only plant standing.
As with Silphium glutinosum, we observed putative hybrids between S. perplexum and other species, especially in roadside populations. By far the most common hybrid was Silphium asteriscus var. asteriscus X S. perplexum (Dallas Co.: A. and S. 8580, VDB; A. and S. 8570, JSU, NY; A. 12156 UNA; Perry Co: A. and S. 8564, AUA, US).
|





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}